

A DnaA-dependent riboswitch for transcription attenuation of the his operon
- PMID: 38817620
- PMCID: PMC10989985
- DOI: 10.1002/mlf2.12075
A DnaA-dependent riboswitch for transcription attenuation of the his operon
Abstract
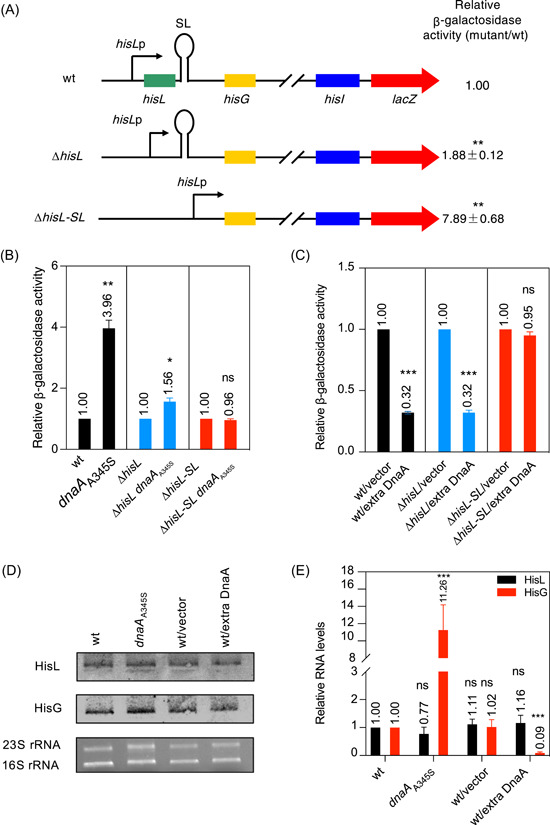
Transcription attenuation in response to the availability of a specific amino acid is believed to be controlled by alternative configurations of RNA secondary structures that lead to the arrest of translation or the release of the arrested ribosome from the leader mRNA molecule. In this study, we first report a possible example of the DnaA-dependent riboswitch for transcription attenuation in Escherichia coli. We show that (i) DnaA regulates the transcription of the structural genes but not that of the leader hisL gene; (ii) DnaA might bind to rDnaA boxes present in the HisL-SL RNA, and subsequently attenuate the transcription of the operon; (iii) the HisL-SL RNA and rDnaA boxes are phylogenetically conserved and evolutionarily important; and (iv) the translating ribosome is required for deattenuation of the his operon, whereas tRNAHis strengthens attenuation. This mechanism seems to be phylogenetically conserved in Gram-negative bacteria and evolutionarily important.
Keywords: DnaA‐dependent; Escherichia coli; his operon; riboswitch; transcription attenuation.
© 2023 The Authors. mLife published by John Wiley & Sons Australia, Ltd. on behalf of Institute of Microbiology, Chinese Academy of Sciences.
Conflict of interest statement
The authors declare no conflict of interests.
Figures



Similar articles
-
Replacement of the Escherichia coli trp operon attenuation control codons alters operon expression.J Mol Biol. 1990 Nov 5;216(1):25-37. doi: 10.1016/S0022-2836(05)80058-0. J Mol Biol. 1990. PMID: 2231731
-
Alternative secondary structures of leader RNAs and the regulation of the trp, phe, his, thr, and leu operons.Proc Natl Acad Sci U S A. 1979 Dec;76(12):6186-90. doi: 10.1073/pnas.76.12.6186. Proc Natl Acad Sci U S A. 1979. PMID: 392514 Free PMC article.
-
The leader mRNA of the histidine attenuator region resembles tRNAHis: possible general regulatory implications.Proc Natl Acad Sci U S A. 1983 Sep;80(17):5240-2. doi: 10.1073/pnas.80.17.5240. Proc Natl Acad Sci U S A. 1983. PMID: 6351055 Free PMC article.
-
Some novel transcription attenuation mechanisms used by bacteria.Biochimie. 1996;78(11-12):1017-24. doi: 10.1016/s0300-9084(97)86725-9. Biochimie. 1996. PMID: 9150880 Review.
-
Transcription attenuation.Biochim Biophys Acta. 2002 Sep 13;1577(2):240-50. doi: 10.1016/s0167-4781(02)00455-4. Biochim Biophys Acta. 2002. PMID: 12213655 Review.
Cited by
-
Read-through transcription of tRNA underlies the cell cycle-dependent dissociation of IHF from the DnaA-inactivating sequence datA.Front Microbiol. 2024 Feb 28;15:1360108. doi: 10.3389/fmicb.2024.1360108. eCollection 2024. Front Microbiol. 2024. PMID: 38505555 Free PMC article.
-
Editorial: Bacterial transcription factors and the cell cycle, volume II.Front Microbiol. 2023 Jul 25;14:1252924. doi: 10.3389/fmicb.2023.1252924. eCollection 2023. Front Microbiol. 2023. PMID: 37560526 Free PMC article. No abstract available.
-
Genome and transcriptomic analysis of the adaptation of Escherichia coli to environmental stresses.Comput Struct Biotechnol J. 2024 May 19;23:2132-2140. doi: 10.1016/j.csbj.2024.05.033. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 38817967 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources