

Differential cell type-specific function of the aryl hydrocarbon receptor and its repressor in diet-induced obesity and fibrosis
- PMID: 38821174
- PMCID: PMC11214421
- DOI: 10.1016/j.molmet.2024.101963
Differential cell type-specific function of the aryl hydrocarbon receptor and its repressor in diet-induced obesity and fibrosis
Abstract
Objective: The aryl hydrocarbon receptor (AhR) is a ligand-activated transcription factor regulating xenobiotic responses as well as physiological metabolism. Dietary AhR ligands activate the AhR signaling axis, whereas AhR activation is negatively regulated by the AhR repressor (AhRR). While AhR-deficient mice are known to be resistant to diet-induced obesity (DIO), the influence of the AhRR on DIO has not been assessed so far.
Methods: In this study, we analyzed AhRR-/- mice and mice with a conditional deletion of either AhRR or AhR in myeloid cells under conditions of DIO and after supplementation of dietary AhR ligands. Moreover, macrophage metabolism was assessed using Seahorse Mito Stress Test and ROS assays as well as transcriptomic analysis.
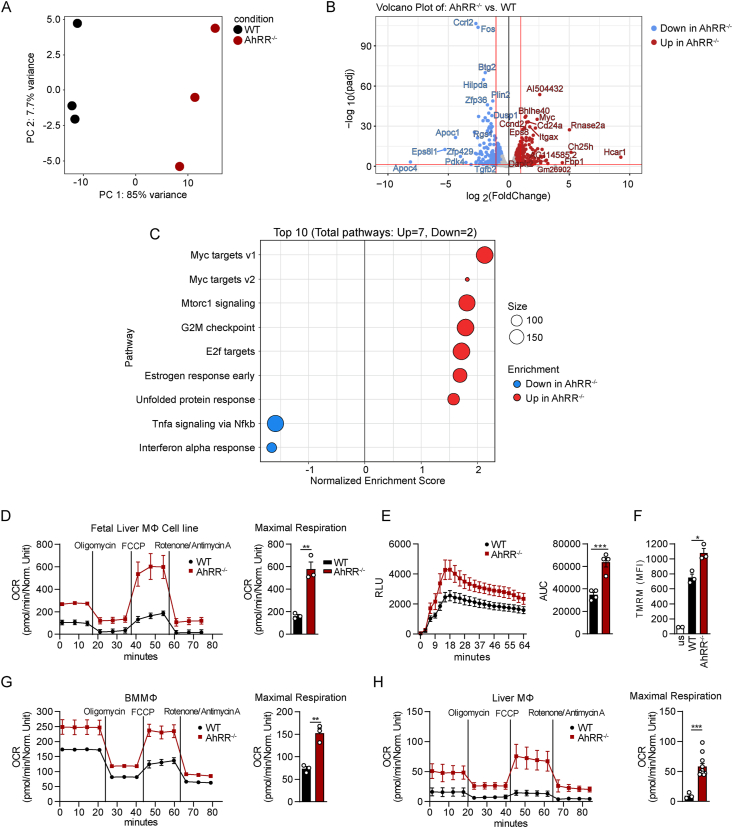
Results: We demonstrate that global AhRR deficiency leads to a robust, but not as profound protection from DIO and hepatosteatosis as AhR deficiency. Under conditions of DIO, AhRR-/- mice did not accumulate TCA cycle intermediates in the circulation in contrast to wild-type (WT) mice, indicating protection from metabolic dysfunction. This effect could be mimicked by dietary supplementation of AhR ligands in WT mice. Because of the predominant expression of the AhRR in myeloid cells, AhRR-deficient macrophages were analyzed for changes in metabolism and showed major metabolic alterations regarding oxidative phosphorylation and mitochondrial activity. Unbiased transcriptomic analysis revealed increased expression of genes involved in de novo lipogenesis and mitochondrial biogenesis. Mice with a genetic deficiency of the AhRR in myeloid cells did not show alterations in weight gain after high fat diet (HFD) but demonstrated ameliorated liver damage compared to control mice. Further, deficiency of the AhR in myeloid cells also did not affect weight gain but led to enhanced liver damage and adipose tissue fibrosis compared to controls.
Conclusions: AhRR-deficient mice are resistant to diet-induced metabolic syndrome. Although conditional ablation of either the AhR or AhRR in myeloid cells did not recapitulate the phenotype of the global knockout, our findings suggest that enhanced AhR signaling in myeloid cells deficient for AhRR protects from diet-induced liver damage and fibrosis, whereas myeloid cell-specific AhR deficiency is detrimental.
Keywords: Collagen deposition; Energy expenditure; Hepatic steatosis; Indole-3-carbinol; Macrophage metabolism; Metabolic dysfunction.
Copyright © 2024 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
