

PDIA3 orchestrates effector T cell program by serving as a chaperone to facilitate the non-canonical nuclear import of STAT1 and PKM2
- PMID: 38822524
- PMCID: PMC11405166
- DOI: 10.1016/j.ymthe.2024.05.038
PDIA3 orchestrates effector T cell program by serving as a chaperone to facilitate the non-canonical nuclear import of STAT1 and PKM2
Erratum in
-
PDIA3 orchestrates effector T cell program by serving as a chaperone to facilitate the non-canonical nuclear import of STAT1 and PKM2.Mol Ther. 2025 Nov 22:S1525-0016(25)00966-9. doi: 10.1016/j.ymthe.2025.11.027. Online ahead of print. Mol Ther. 2025. PMID: 41275860 No abstract available.
Abstract
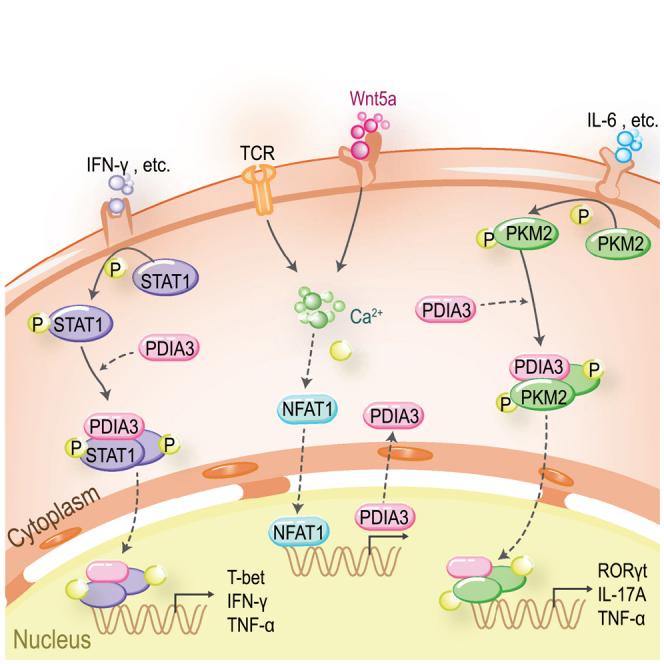
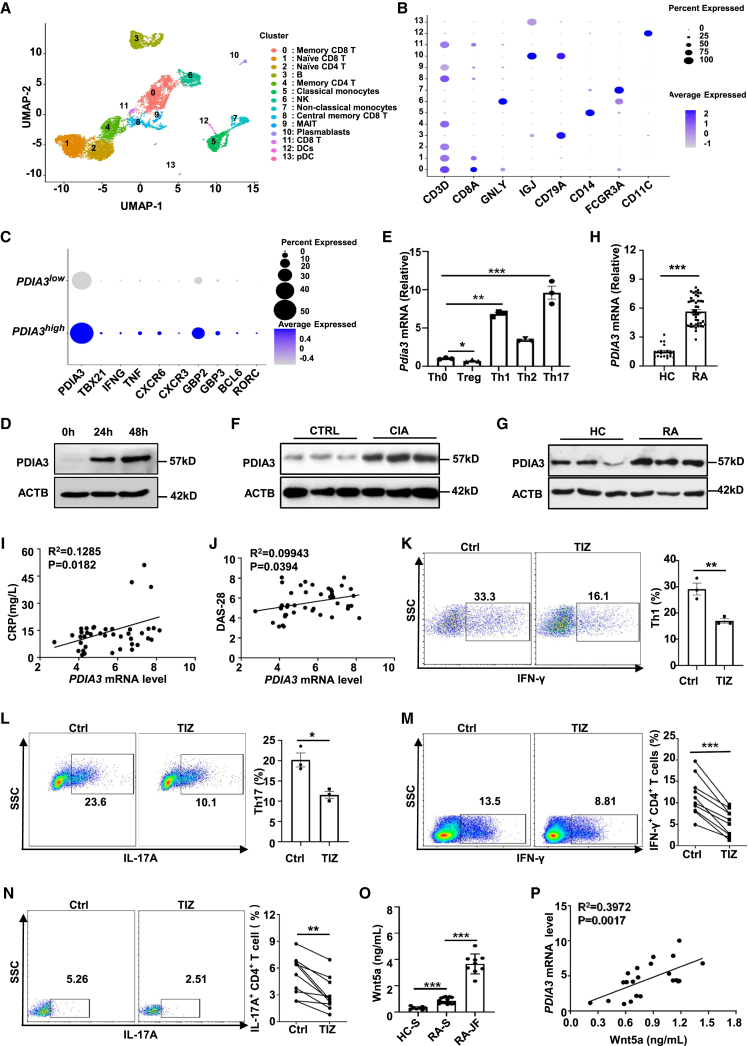
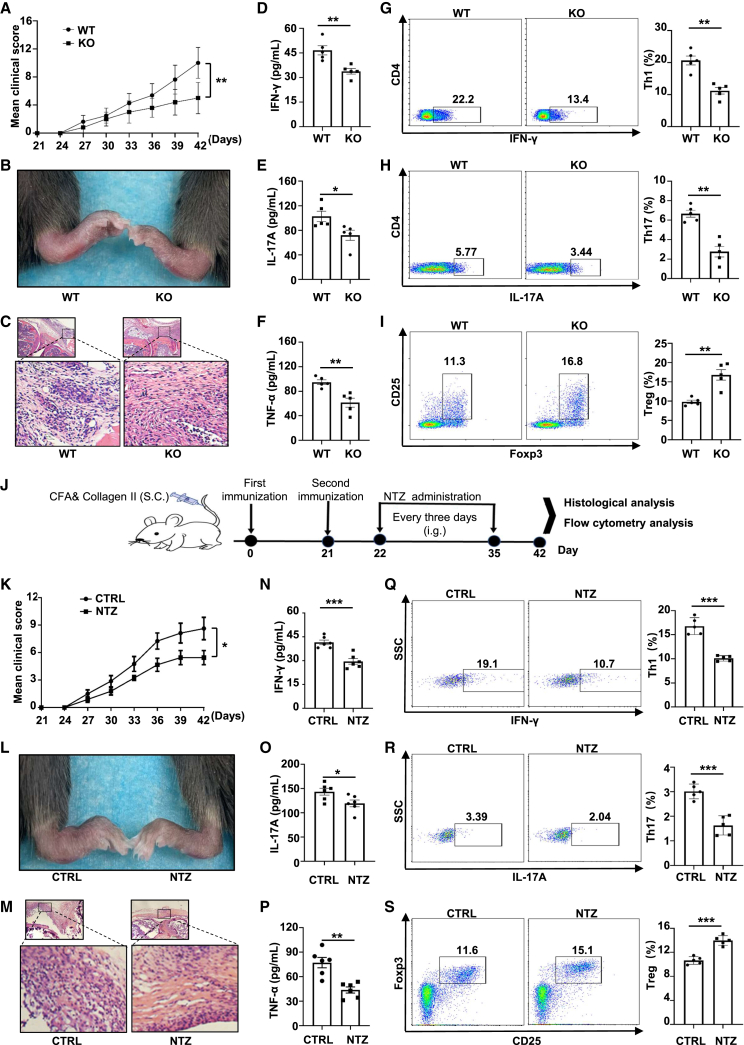
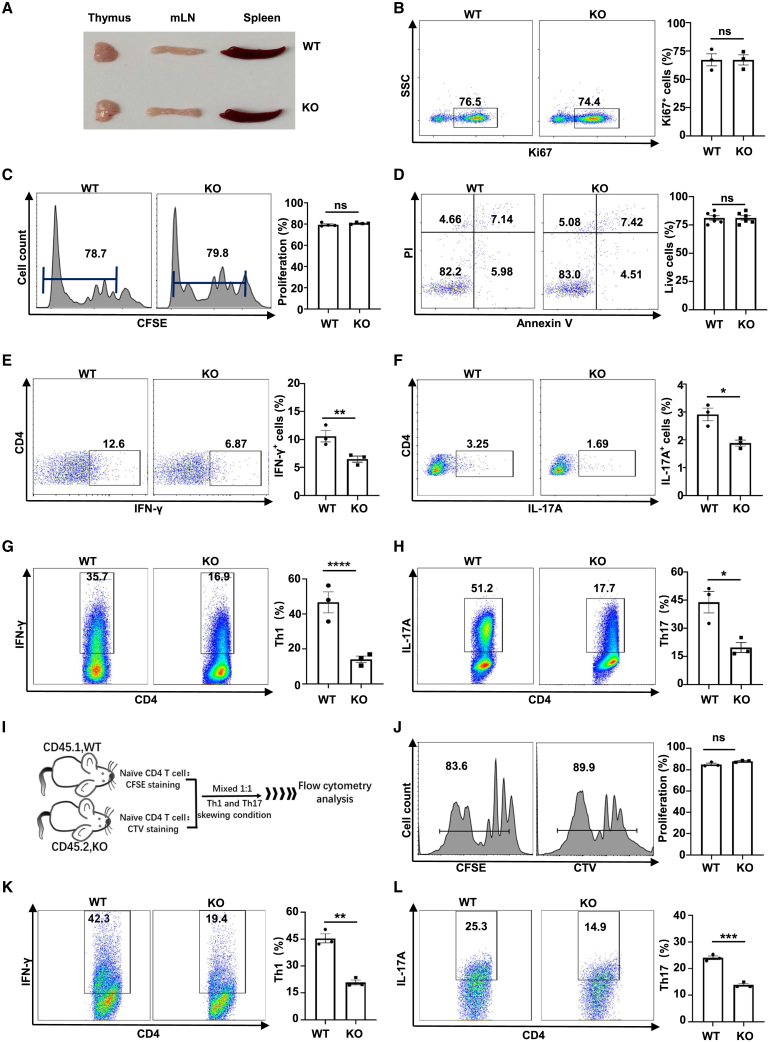
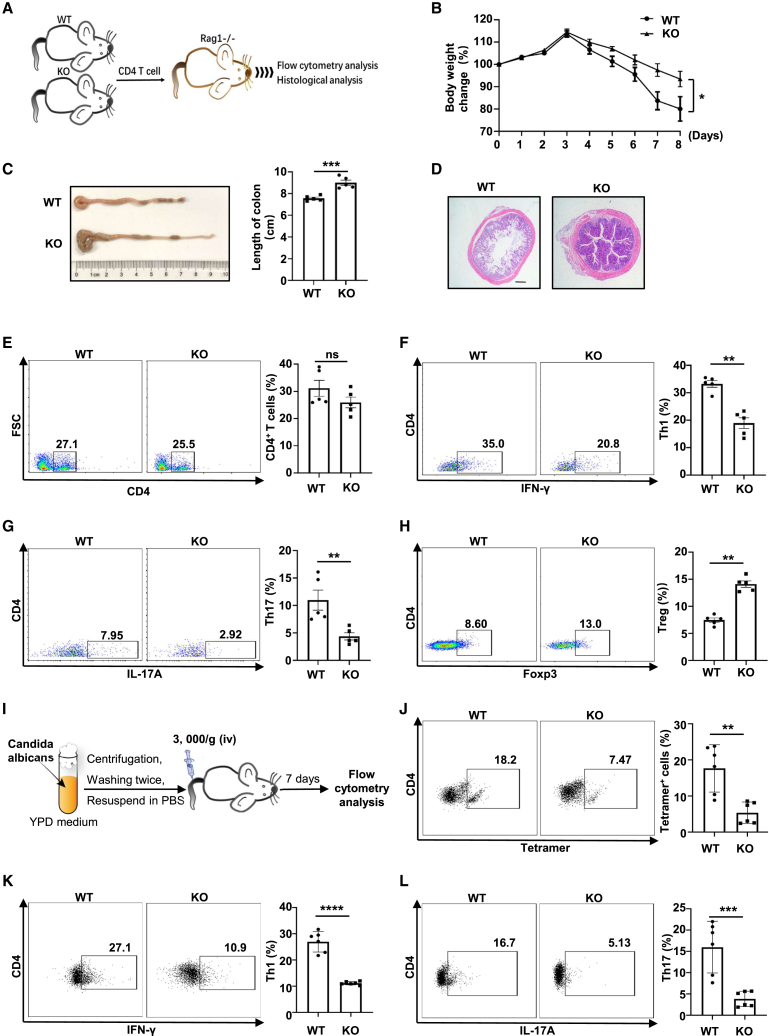
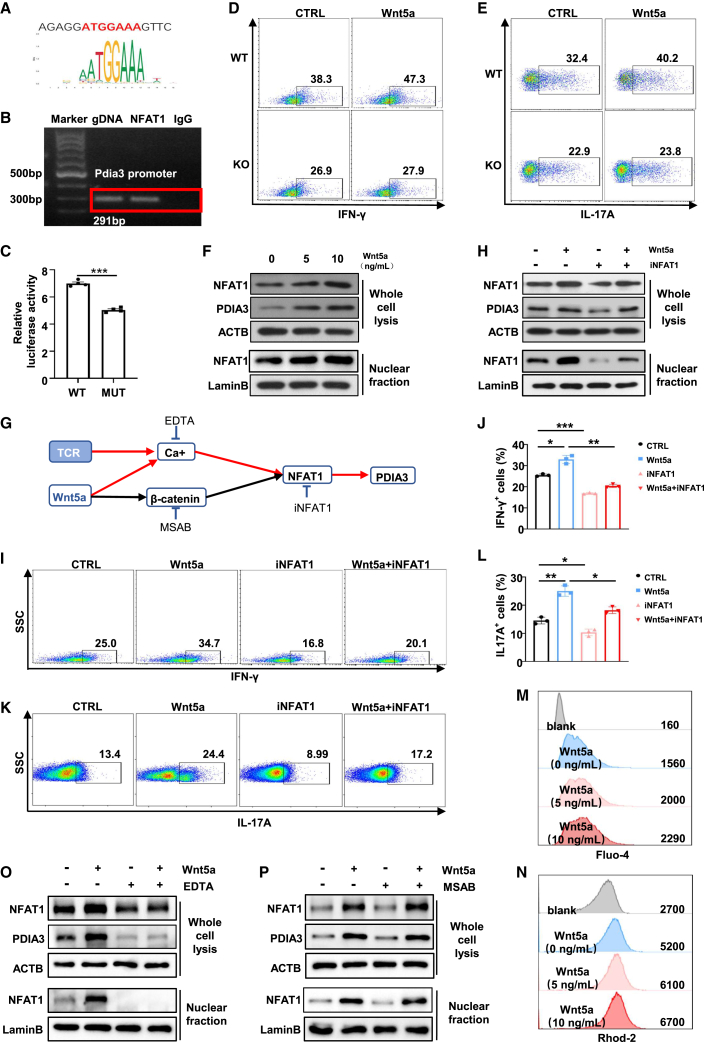
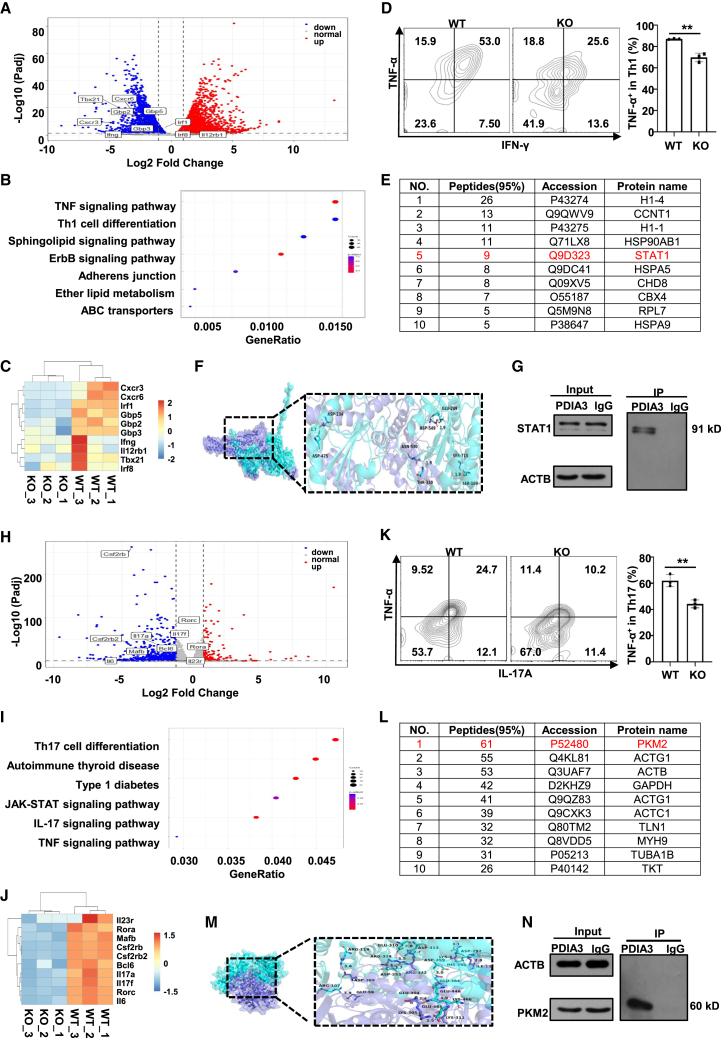
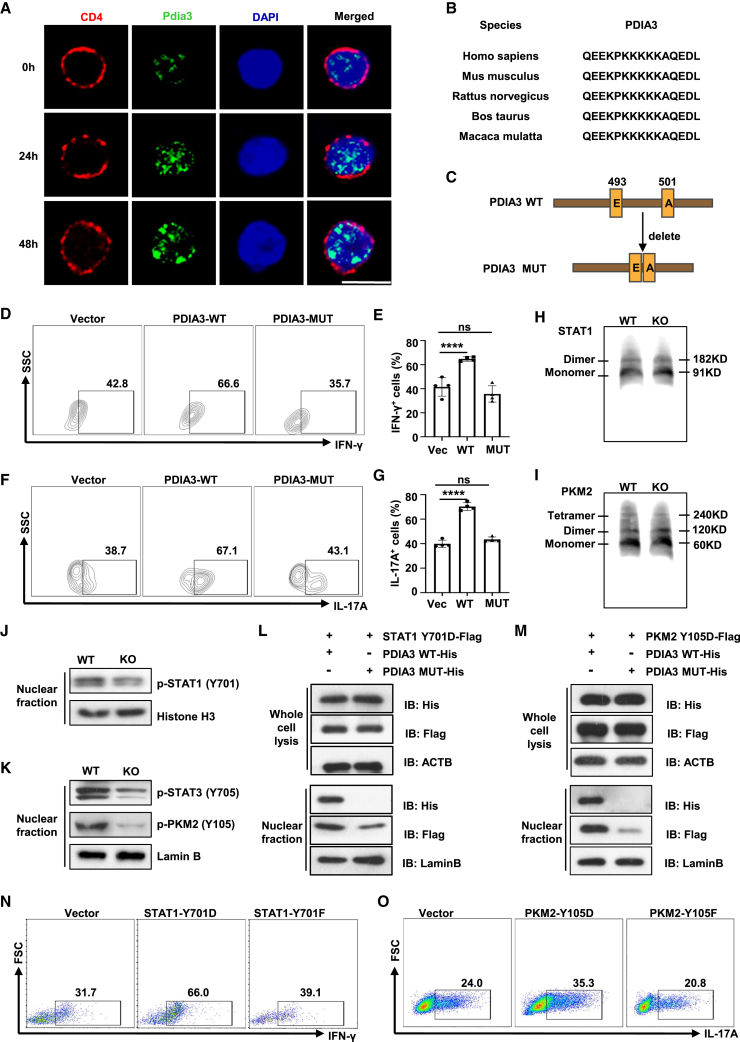
Dysregulated T cell activation underpins the immunopathology of rheumatoid arthritis (RA), yet the machineries that orchestrate T cell effector program remain incompletely understood. Herein, we leveraged bulk and single-cell RNA sequencing data from RA patients and validated protein disulfide isomerase family A member 3 (PDIA3) as a potential therapeutic target. PDIA3 is remarkably upregulated in pathogenic CD4 T cells derived from RA patients and positively correlates with C-reactive protein level and disease activity score 28. Pharmacological inhibition or genetic ablation of PDIA3 alleviates RA-associated articular pathology and autoimmune responses. Mechanistically, T cell receptor signaling triggers intracellular calcium flux to activate NFAT1, a process that is further potentiated by Wnt5a under RA settings. Activated NFAT1 then directly binds to the Pdia3 promoter to enhance the expression of PDIA3, which complexes with STAT1 or PKM2 to facilitate their nuclear import for transcribing T helper 1 (Th1) and Th17 lineage-related genes, respectively. This non-canonical regulatory mechanism likely occurs under pathological conditions, as PDIA3 could only be highly induced following aberrant external stimuli. Together, our data support that targeting PDIA3 is a vital strategy to mitigate autoimmune diseases, such as RA, in clinical settings.
Keywords: PDIA3; effector T cell program; non-canonical function; nuclear transport; rheumatoid arthritis.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare that they have no conflict of interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
