

Lipid switches in the immunological synapse
- PMID: 38823638
- PMCID: PMC11259711
- DOI: 10.1016/j.jbc.2024.107428
Lipid switches in the immunological synapse
Abstract
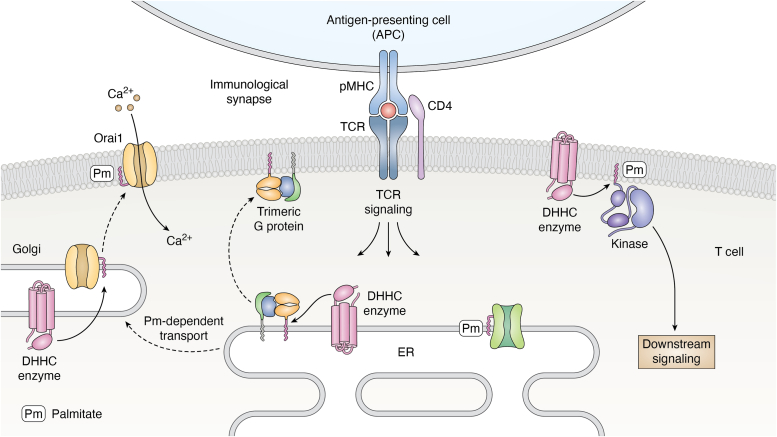
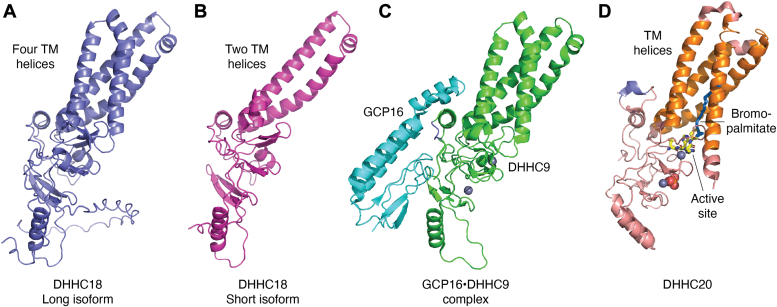
Adaptive immune responses comprise the activation of T cells by peptide antigens that are presented by proteins of the Major Histocompatibility Complex (MHC) on the surface of an antigen-presenting cell. As a consequence of the T cell receptor interacting productively with a certain peptide-MHC complex, a specialized cell-cell junction known as the immunological synapse forms and is accompanied by changes in the spatiotemporal patterning and function of intracellular signaling molecules. Key modifications occurring at the cytoplasmic leaflet of the plasma and internal membranes in activated T cells comprise lipid switches that affect the binding and distribution of proteins within or near the lipid bilayer. Here, we describe two major classes of lipid switches that act at this critical water/membrane interface. Phosphoinositides are derived from phosphatidylinositol, an amphiphilic molecule that contains two fatty acid chains and a phosphate group that bridges the glycerol backbone to the carbohydrate inositol. The inositol ring can be variably (de-)phosphorylated by dedicated kinases and phosphatases, thereby creating phosphoinositide signatures that define the composition and properties of signaling molecules, molecular complexes, or whole organelles. Palmitoylation refers to the reversible attachment of the fatty acid palmitate to a substrate protein's cysteine residue. DHHC enzymes, named after the four conserved amino acids in their active site, catalyze this post-translational modification and thereby change the distribution of proteins at, between, and within membranes. T cells utilize these two types of molecular switches to adjust their properties to an activation process that requires changes in motility, transport, secretion, and gene expression.
Keywords: DHHC enzymes; S-acylation; T cell; immunological synapse; palmitoylation; phosphoinositides.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures


References
-
- Grakoui A., Bromley S.K., Sumen C., Davis M.M., Shaw A.S., Allen P.M., et al. The immunological synapse: a molecular machine controlling T cell activation. Science. 1999;285:221–227. - PubMed
-
- Dustin M.L., Choudhuri K. Signaling and polarized communication across the T cell immunological synapse. Annu. Rev. Cell Dev. Biol. 2016;32:303–325. - PubMed
-
- Wu W., Shi X., Xu C. Regulation of T cell signalling by membrane lipids. Nat. Rev. Immunol. 2016;16:690–701. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
