

This is a preprint.
Granzyme K drives a newly-intentified pathway of complement activation
- PMID: 38826230
- PMCID: PMC11142156
- DOI: 10.1101/2024.05.22.595315
Granzyme K drives a newly-intentified pathway of complement activation
Update in
-
Granzyme K activates the entire complement cascade.Nature. 2025 May;641(8061):211-221. doi: 10.1038/s41586-025-08713-9. Epub 2025 Feb 6. Nature. 2025. PMID: 39914456 Free PMC article.
Abstract
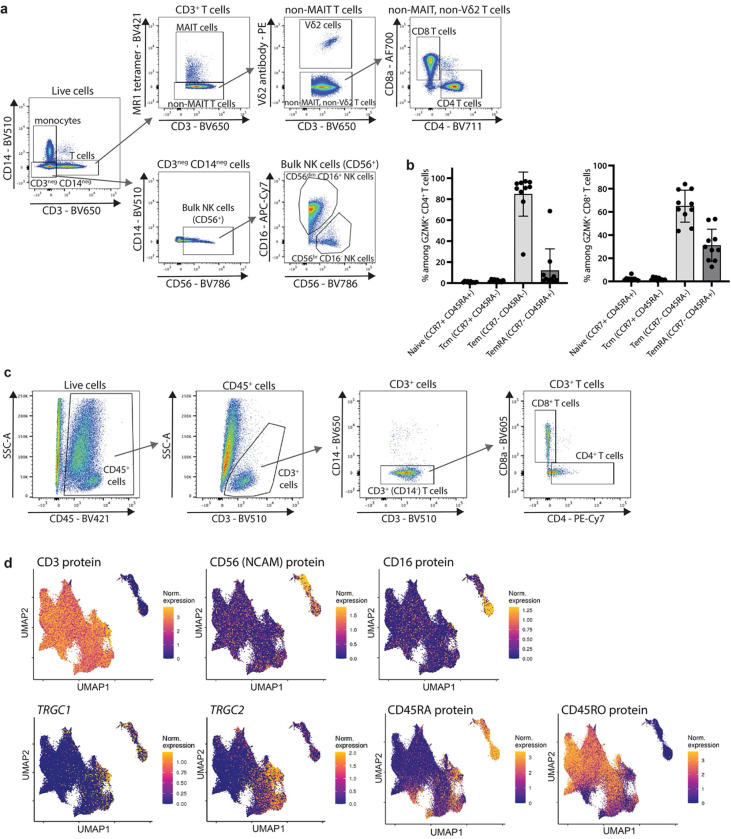
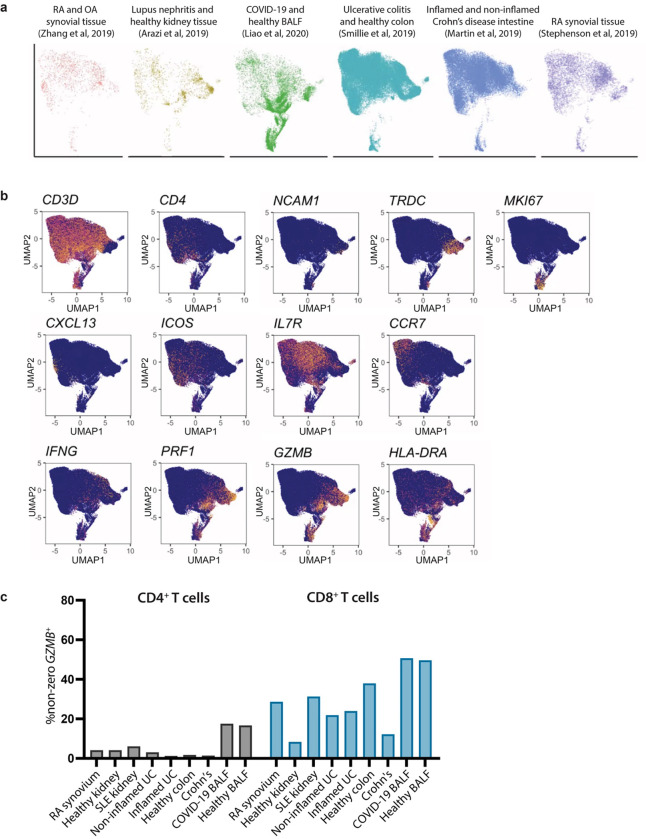
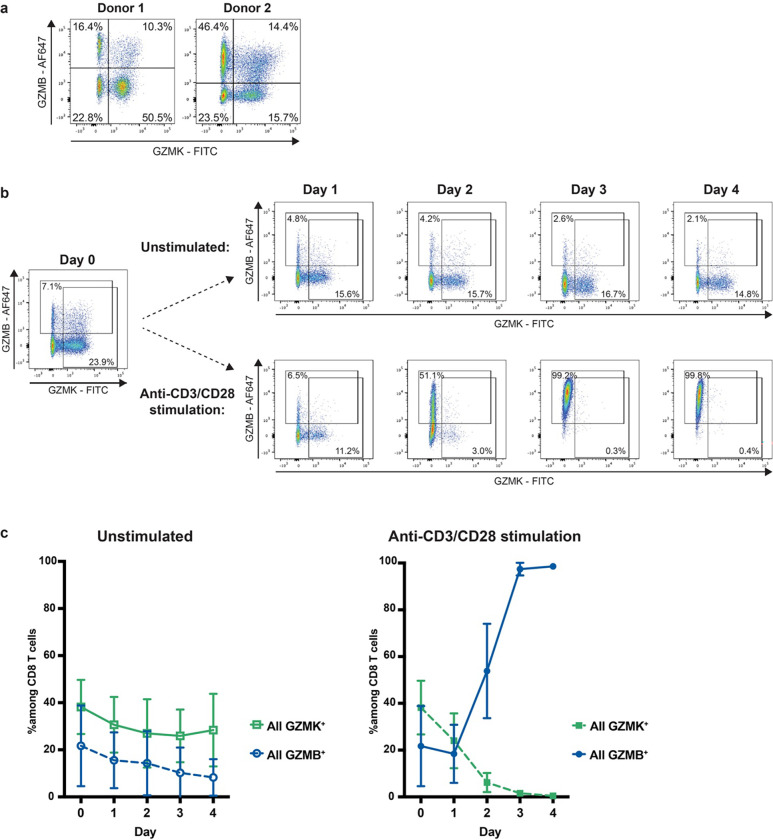
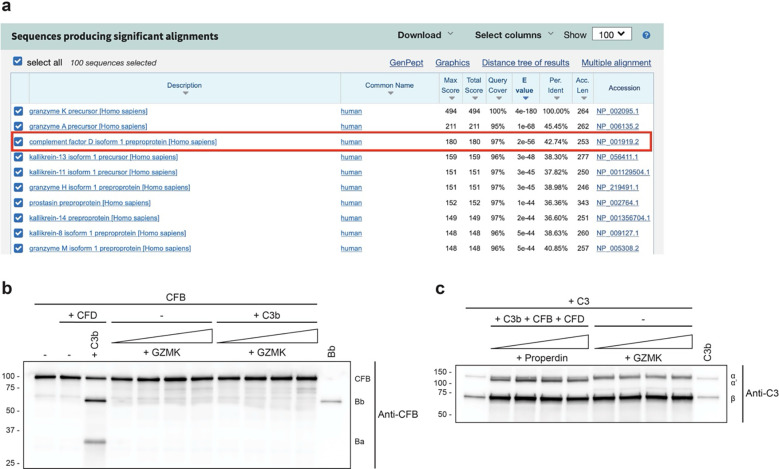
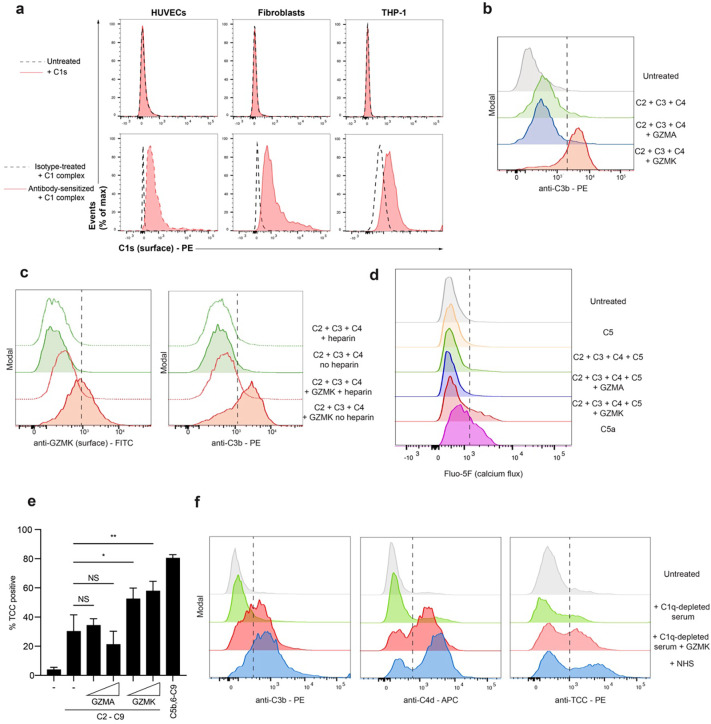
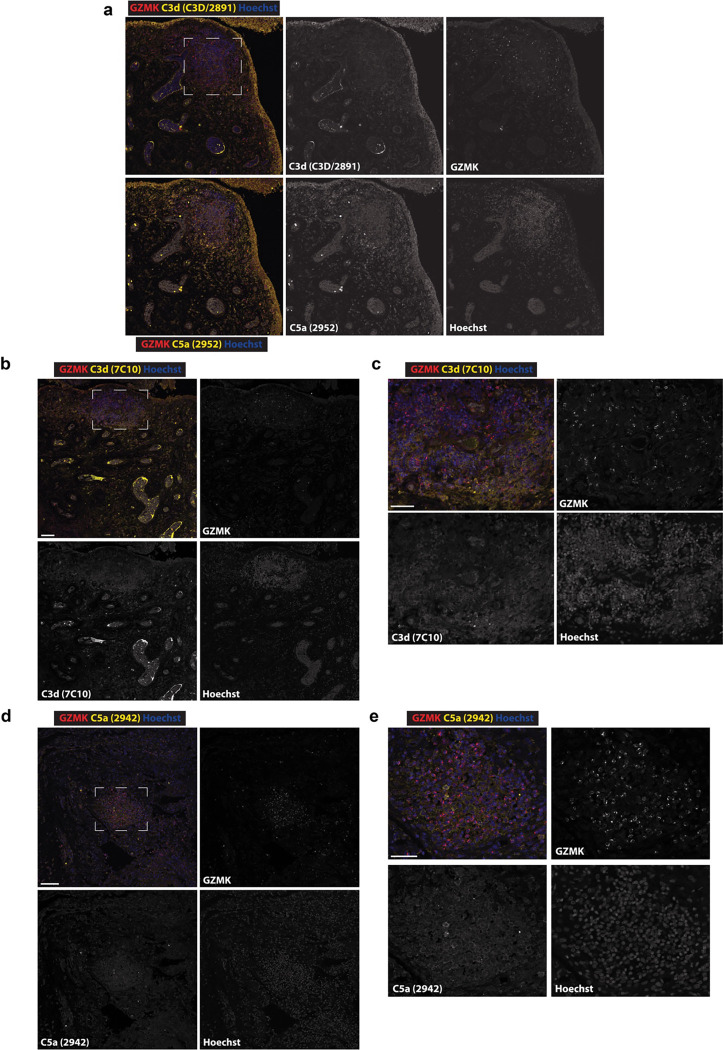
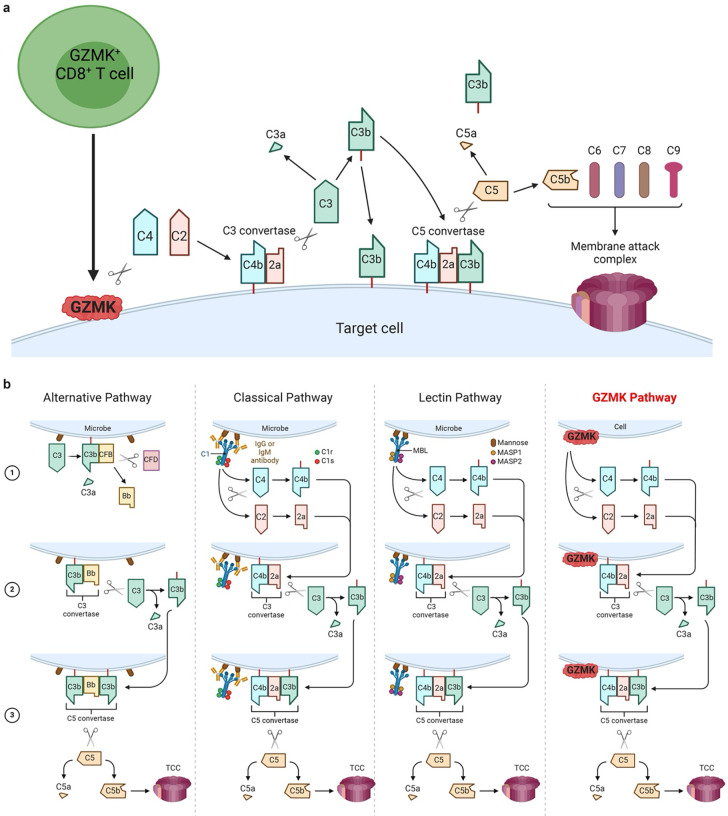
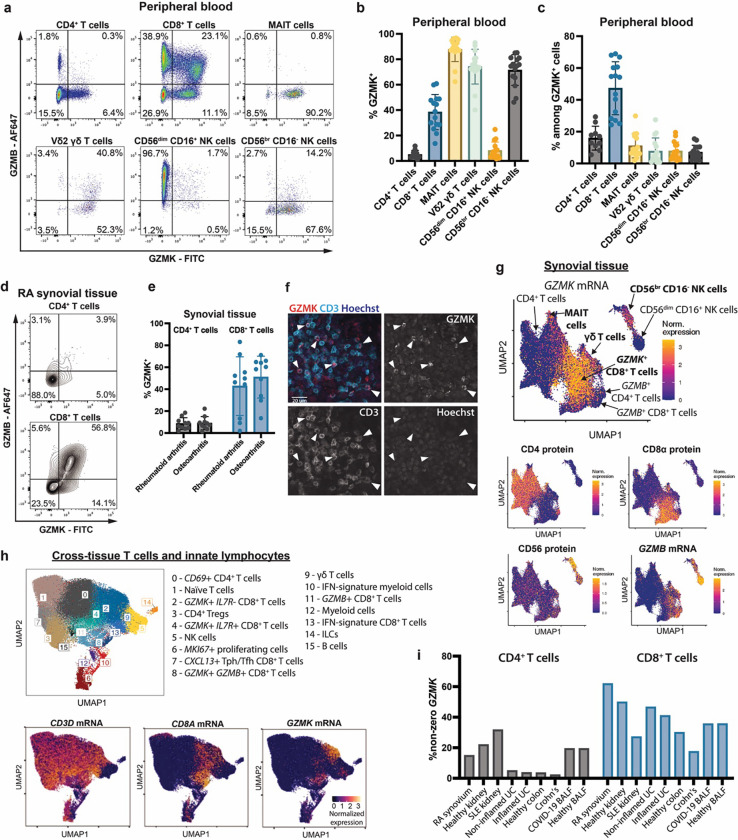
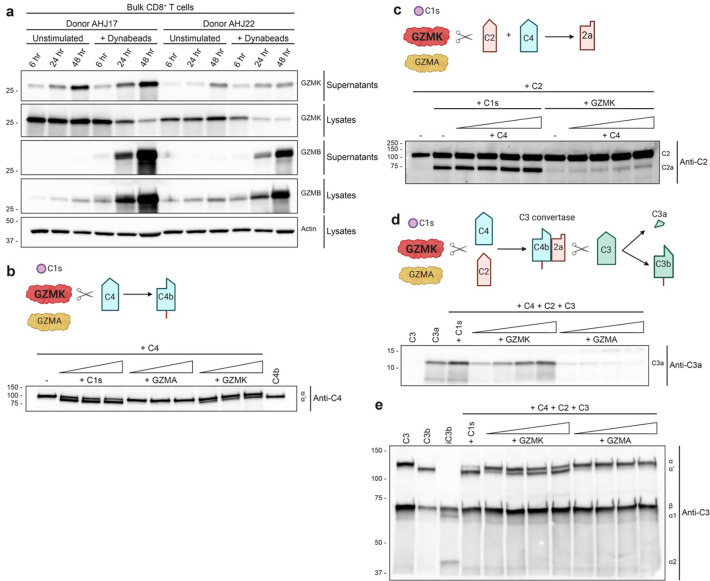
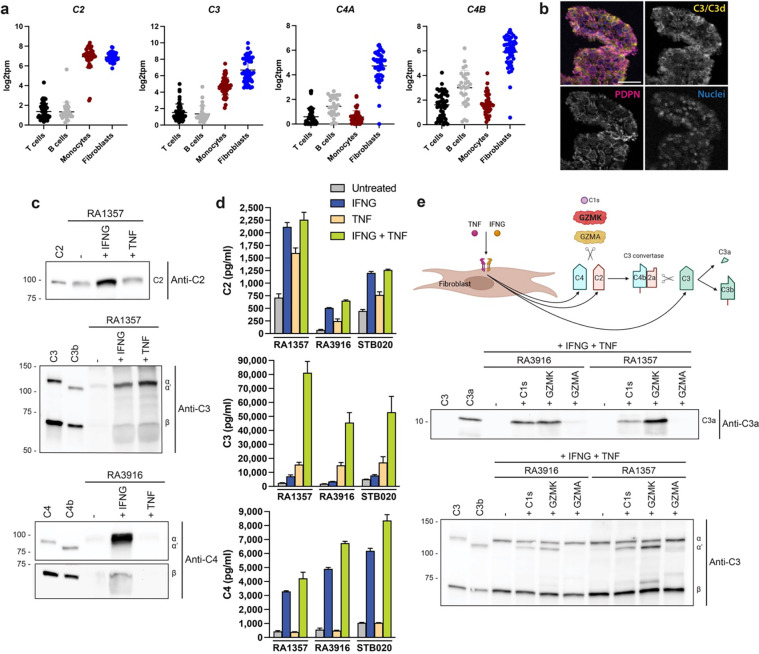
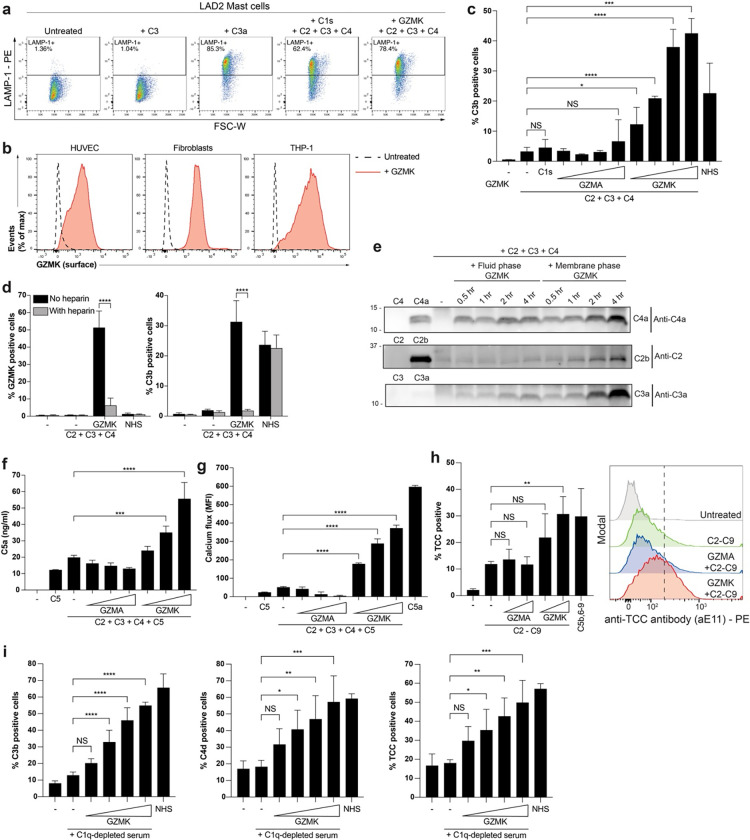
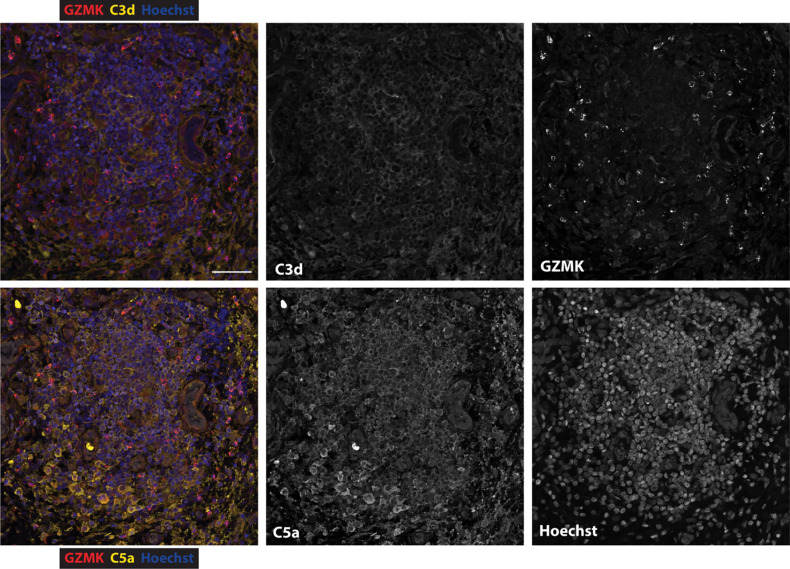
Granzymes are a family of serine proteases mainly expressed by CD8+ T cells, natural killer cells, and innate-like lymphocytes1,2. Although their major role is thought to be the induction of cell death in virally infected and tumor cells, accumulating evidence suggests some granzymes can regulate inflammation by acting on extracellular substrates2. Recently, we found that the majority of tissue CD8+ T cells in rheumatoid arthritis (RA) synovium, inflammatory bowel disease and other inflamed organs express granzyme K (GZMK)3, a tryptase-like protease with poorly defined function. Here, we show that GZMK can activate the complement cascade by cleaving C2 and C4. The nascent C4b and C2a fragments form a C3 convertase that cleaves C3, allowing further assembly of a C5 convertase that cleaves C5. The resulting convertases trigger every major event in the complement cascade, generating the anaphylatoxins C3a and C5a, the opsonins C4b and C3b, and the membrane attack complex. In RA synovium, GZMK is enriched in areas with abundant complement activation, and fibroblasts are the major producers of complement C2, C3, and C4 that serve as targets for GZMK-mediated complement activation. Our findings describe a previously unidentified pathway of complement activation that is entirely driven by lymphocyte-derived GZMK and proceeds independently of the classical, lectin, or alternative pathways. Given the widespread abundance of GZMK-expressing T cells in tissues in chronic inflammatory diseases and infection, GZMK-mediated complement activation is likely to be an important contributor to tissue inflammation in multiple disease contexts.
Figures
References
-
- Anthony D. A., Andrews D. M., Watt S. V., Trapani J. A. & Smyth M. J. Functional dissection of the granzyme family: cell death and inflammation. Immunol. Rev. 235, 73–92 (2010). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous