

This is a preprint.
Differential TLR-ERK1/2 activity promotes viral ssRNA and dsRNA mimic-induced dysregulated immunity in macrophages
- PMID: 38826464
- PMCID: PMC11142249
- DOI: 10.1101/2024.05.24.595760
Differential TLR-ERK1/2 activity promotes viral ssRNA and dsRNA mimic-induced dysregulated immunity in macrophages
Update in
-
Differential TLR-ERK1/2 Activity Promotes Viral ssRNA and dsRNA Mimic-Induced Dysregulated Immunity in Macrophages.Pathogens. 2024 Nov 23;13(12):1033. doi: 10.3390/pathogens13121033. Pathogens. 2024. PMID: 39770293 Free PMC article.
Abstract
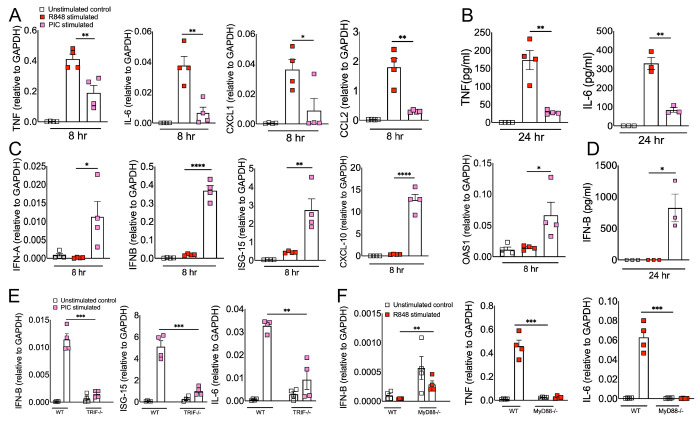
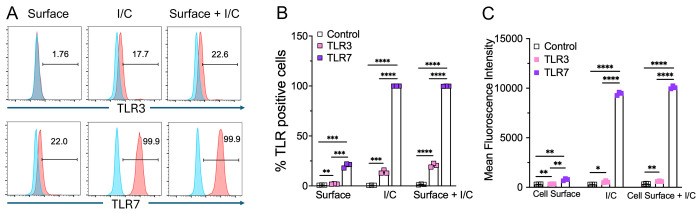
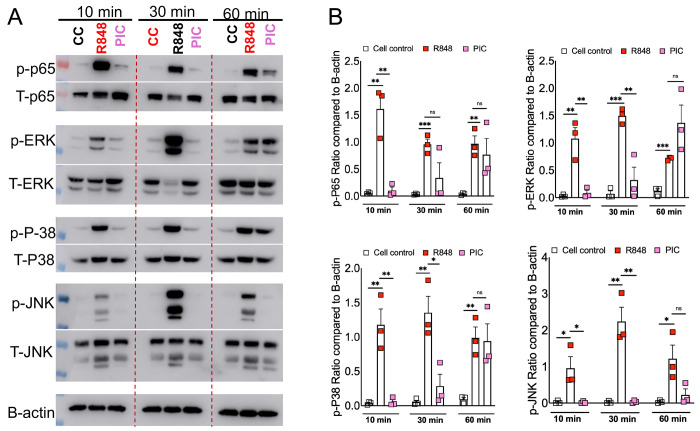
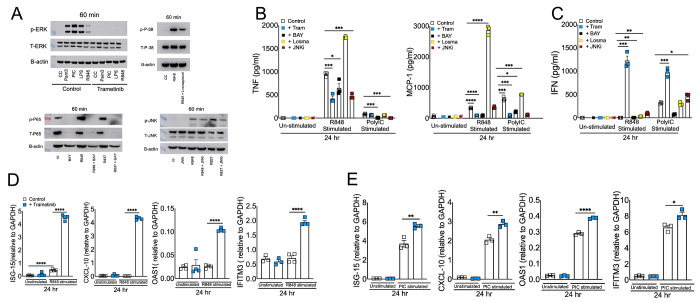
RNA virus induced excessive inflammation and impaired antiviral interferon (IFN-I) responses are associated with severe disease. This innate immune response, also referred to as 'dysregulated immunity,' is caused by viral single-stranded RNA (ssRNA) and double-stranded-RNA (dsRNA) mediated exuberant inflammation and viral protein-induced IFN antagonism. However, key host factors and the underlying mechanism driving viral RNA-mediated dysregulated immunity are poorly defined. Here, using viral ssRNA and dsRNA mimics, which activate toll-like receptor 7 (TLR7) and TLR3, respectively, we evaluated the role of viral RNAs in causing dysregulated immunity. We show that murine bone marrow-derived macrophages (BMDMs) stimulated with TLR3 and TLR7 agonists induce differential inflammatory and antiviral cytokine response. TLR7 activation triggered a robust inflammatory cytokine/chemokine induction compared to TLR3 activation, whereas TLR3 stimulation induced significantly increased IFN/IFN stimulated gene (ISG) response relative to TLR7 activation. To define the mechanistic basis for dysregulated immunity, we examined cell-surface and endosomal TLR levels and downstream mitogen-activated protein kinase (MAPK) and nuclear factor kappa B (NF-kB) activation. We identified a significantly higher cell-surface and endosomal TLR7 expression compared to TLR3, which further correlated with early and robust MAPK (pERK1/2 and p-P38) and NF-kB activation in TLR7-stimulated macrophages. Furthermore, blocking EKR1/2, p38, and NF-kB activity reduced TLR3/7-induced inflammatory cytokine/chemokine levels, whereas only ERK1/2 inhibition enhanced viral RNA-mimic-induced IFN/ISG responses. Collectively, our results illustrate that high cell surface and endosomal TLR7 expression and robust ERK1/2 activation drive viral ssRNA mimic-induced excessive inflammatory and reduced IFN/ISG responses, and blocking ERK1/2 activity would mitigate viral-RNA/TLR-induced dysregulated immunity.
Keywords: ERK1/2; SARS-COV-2; TLRs; inflammation; interferon; macrophages.
Figures
References
-
- Wang LF, Eaton BT. Bats, Civets and the Emergence of SARS. In: Childs JE, Mackenzie JS, Richt JA, eds. Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission. Springer; 2007:325–344. doi: 10.1007/978-3-540-70962-6_13 - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous