

Epistasis arises from shifting the rate-limiting step during enzyme evolution of a β-lactamase
- PMID: 38828429
- PMCID: PMC11136654
- DOI: 10.1038/s41929-024-01117-4
Epistasis arises from shifting the rate-limiting step during enzyme evolution of a β-lactamase
Abstract
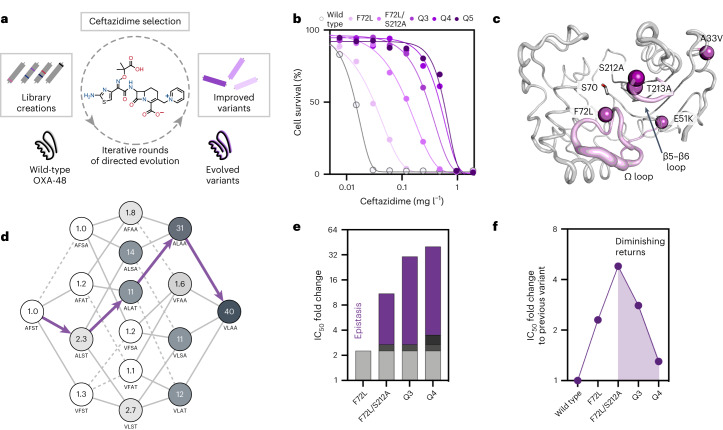
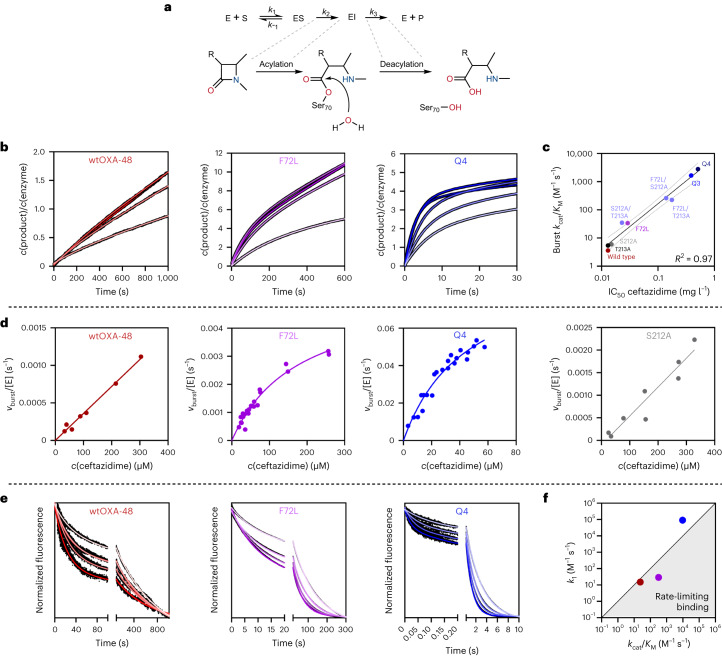
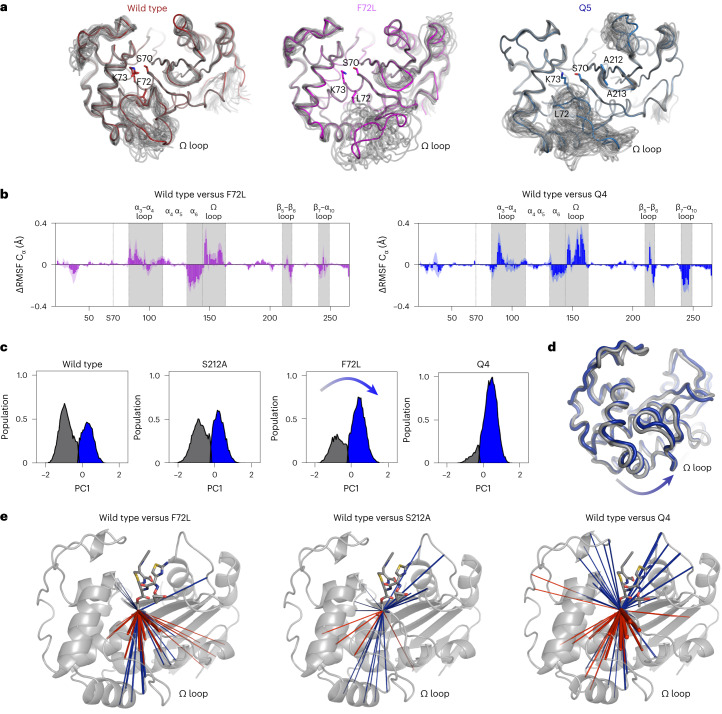
Epistasis, the non-additive effect of mutations, can provide combinatorial improvements to enzyme activity that substantially exceed the gains from individual mutations. Yet the molecular mechanisms of epistasis remain elusive, undermining our ability to predict pathogen evolution and engineer biocatalysts. Here we reveal how directed evolution of a β-lactamase yielded highly epistatic activity enhancements. Evolution selected four mutations that increase antibiotic resistance 40-fold, despite their marginal individual effects (≤2-fold). Synergistic improvements coincided with the introduction of super-stochiometric burst kinetics, indicating that epistasis is rooted in the enzyme's conformational dynamics. Our analysis reveals that epistasis stemmed from distinct effects of each mutation on the catalytic cycle. The initial mutation increased protein flexibility and accelerated substrate binding, which is rate-limiting in the wild-type enzyme. Subsequent mutations predominantly boosted the chemical steps by fine-tuning substrate interactions. Our work identifies an overlooked cause for epistasis: changing the rate-limiting step can result in substantial synergy that boosts enzyme activity.
Keywords: Biocatalysis; Enzyme mechanisms; Hydrolases; Molecular evolution.
© The Author(s) 2024, corrected publication 2024.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures
Similar articles
-
Negative Epistasis and Evolvability in TEM-1 β-Lactamase--The Thin Line between an Enzyme's Conformational Freedom and Disorder.J Mol Biol. 2015 Jul 17;427(14):2396-409. doi: 10.1016/j.jmb.2015.05.011. Epub 2015 May 22. J Mol Biol. 2015. PMID: 26004540 Free PMC article.
-
Patterns of Epistasis between beneficial mutations in an antibiotic resistance gene.Mol Biol Evol. 2013 Aug;30(8):1779-87. doi: 10.1093/molbev/mst096. Epub 2013 May 15. Mol Biol Evol. 2013. PMID: 23676768 Free PMC article.
-
Network of epistatic interactions in an enzyme active site revealed by large-scale deep mutational scanning.Proc Natl Acad Sci U S A. 2024 Mar 19;121(12):e2313513121. doi: 10.1073/pnas.2313513121. Epub 2024 Mar 14. Proc Natl Acad Sci U S A. 2024. PMID: 38483989 Free PMC article.
-
Quantitative interpretations of double mutations of enzymes.Arch Biochem Biophys. 1992 May 1;294(2):327-40. doi: 10.1016/0003-9861(92)90692-p. Arch Biochem Biophys. 1992. PMID: 1567189 Review.
-
Deleterious mutations, variable epistatic interactions, and the evolution of recombination.Theor Popul Biol. 1997 Apr;51(2):134-47. doi: 10.1006/tpbi.1997.1301. Theor Popul Biol. 1997. PMID: 9169238 Review.
Cited by
-
Dynamical responses predict a distal site that modulates activity in an antibiotic resistance enzyme.Chem Sci. 2024 Sep 30;15(41):17232-44. doi: 10.1039/d4sc03295k. Online ahead of print. Chem Sci. 2024. PMID: 39364073 Free PMC article.
-
Electric Fields Are a Key Determinant of Carbapenemase Activity in Class A β-Lactamases.ACS Catal. 2024 Apr 23;14(9):7166-7172. doi: 10.1021/acscatal.3c05302. eCollection 2024 May 3. ACS Catal. 2024. PMID: 38721371 Free PMC article.
-
Structural comparison of substrate-binding pockets of serine β-lactamases in classes A, C, and D.J Enzyme Inhib Med Chem. 2025 Dec;40(1):2435365. doi: 10.1080/14756366.2024.2435365. Epub 2024 Dec 23. J Enzyme Inhib Med Chem. 2025. PMID: 39714271 Free PMC article. Review.
-
Active learning-assisted directed evolution.Nat Commun. 2025 Jan 16;16(1):714. doi: 10.1038/s41467-025-55987-8. Nat Commun. 2025. PMID: 39821082 Free PMC article.
-
Effect of Mutations on the Evolution of Extended Spectrum β-lactamases (ESBL).Protein J. 2025 Aug 19. doi: 10.1007/s10930-025-10284-7. Online ahead of print. Protein J. 2025. PMID: 40830675 Review.
References
-
- Buda, K., Miton, C. M. & Tokuriki, N. Higher-order epistasis creates idiosyncrasy, confounding predictions in protein evolution. Nat. Commun.10.1038/s41467-023-44333-5 (2023).
Grants and funding
LinkOut - more resources
Full Text Sources