

Anisotropic Network Analysis of Open/Closed HCN4 Channel Advocates Asymmetric Subunit Cooperativity in cAMP Modulation of Gating
- PMID: 38830626
- PMCID: PMC11203669
- DOI: 10.1021/acs.jcim.4c00360
Anisotropic Network Analysis of Open/Closed HCN4 Channel Advocates Asymmetric Subunit Cooperativity in cAMP Modulation of Gating
Abstract
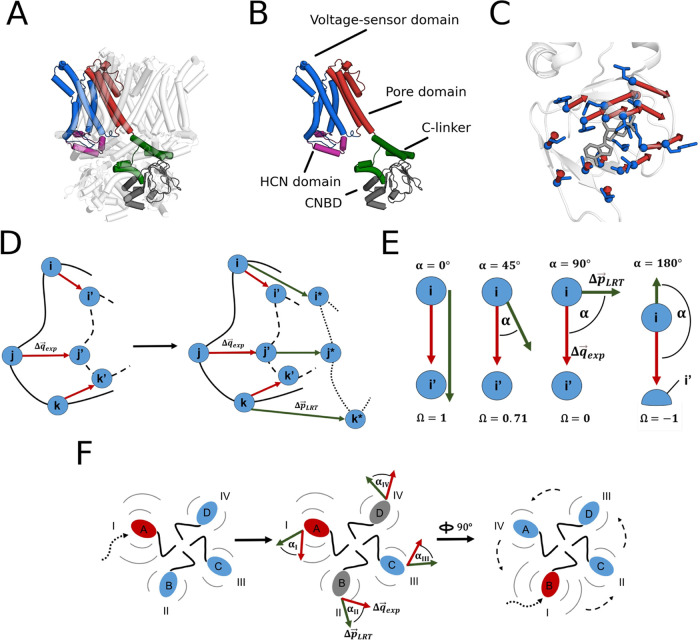
Hyperpolarization-activated cyclic nucleotide-modulated (HCN) channels are opened in an allosteric manner by membrane hyperpolarization and cyclic nucleotides such as cAMP. Because of conflicting reports from experimental studies on whether cAMP binding to the four available binding sites in the channel tetramer operates cooperatively in gating, we employ here a computational approach as a promising route to examine ligand-induced conformational changes after binding to individual sites. By combining an elastic network model (ENM) with linear response theory (LRT) for modeling the apo-holo transition of the cyclic nucleotide-binding domain (CNBD) in HCN channels, we observe a distinct pattern of cooperativity matching the "positive-negative-positive" cooperativity reported from functional studies. This cooperativity pattern is highly conserved among HCN subtypes (HCN4, HCN1), but only to a lesser extent visible in structurally related channels, which are only gated by voltage (KAT1) or cyclic nucleotides (TAX4). This suggests an inherent cooperativity between subunits in HCN channels as part of a ligand-triggered gating mechanism in these channels.
Conflict of interest statement
The authors declare no competing financial interest.
Figures



Similar articles
-
The structure of the apo cAMP-binding domain of HCN4 - a stepping stone toward understanding the cAMP-dependent modulation of the hyperpolarization-activated cyclic-nucleotide-gated ion channels.FEBS J. 2018 Jun;285(12):2182-2192. doi: 10.1111/febs.14408. Epub 2018 Mar 14. FEBS J. 2018. PMID: 29444387 Review.
-
The HCN domain is required for HCN channel cell-surface expression and couples voltage- and cAMP-dependent gating mechanisms.J Biol Chem. 2020 Jun 12;295(24):8164-8173. doi: 10.1074/jbc.RA120.013281. Epub 2020 Apr 27. J Biol Chem. 2020. PMID: 32341127 Free PMC article.
-
A mechanism for the auto-inhibition of hyperpolarization-activated cyclic nucleotide-gated (HCN) channel opening and its relief by cAMP.J Biol Chem. 2014 Aug 8;289(32):22205-20. doi: 10.1074/jbc.M114.572164. Epub 2014 May 30. J Biol Chem. 2014. PMID: 24878962 Free PMC article.
-
The HCN domain couples voltage gating and cAMP response in hyperpolarization-activated cyclic nucleotide-gated channels.Elife. 2019 Nov 26;8:e49672. doi: 10.7554/eLife.49672. Elife. 2019. PMID: 31769408 Free PMC article.
-
Structural and functional approaches to studying cAMP regulation of HCN channels.Biochem Soc Trans. 2021 Dec 17;49(6):2573-2579. doi: 10.1042/BST20210290. Biochem Soc Trans. 2021. PMID: 34812892 Review.
References
-
- Saponaro A.; Bauer D.; Giese M. H.; Swuec P.; Porro A.; Gasparri F.; Sharifzadeh A. S.; Chaves-Sanjuan A.; Alberio L.; Parisi G.; Cerutti G.; Clarke O. B.; Hamacher K.; Colecraft H. M.; Mancia F.; Hendrickson W. A.; Siegelbaum S. A.; DiFrancesco D.; Bolognesi M.; Thiel G.; Santoro B.; Moroni A. Gating Movements and Ion Permeation in HCN4 Pacemaker Channels. Mol. Cell 2021, 81 (14), 2929–2943. 10.1016/j.molcel.2021.05.033. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
