

The human alpha7 nicotinic acetylcholine receptor is a host target for the rabies virus glycoprotein
- PMID: 38836054
- PMCID: PMC11148329
- DOI: 10.3389/fcimb.2024.1394713
The human alpha7 nicotinic acetylcholine receptor is a host target for the rabies virus glycoprotein
Abstract
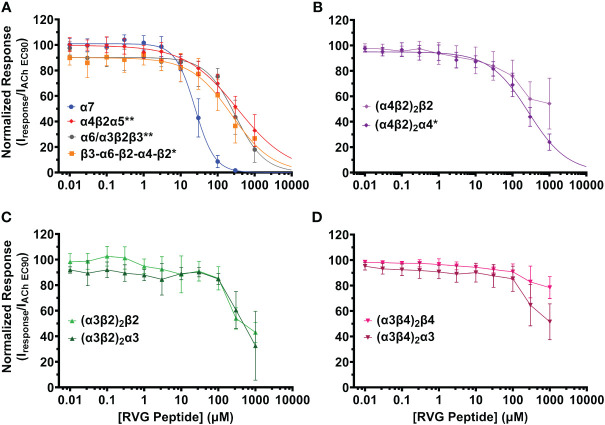
The rabies virus enters the nervous system by interacting with several molecular targets on host cells to modify behavior and trigger receptor-mediated endocytosis of the virion by poorly understood mechanisms. The rabies virus glycoprotein (RVG) interacts with the muscle acetylcholine receptor and the neuronal α4β2 subtype of the nicotinic acetylcholine receptor (nAChR) family by the putative neurotoxin-like motif. Given that the neurotoxin-like motif is highly homologous to the α7 nAChR subtype selective snake toxin α-bungarotoxin (αBTX), other nAChR subtypes are likely involved. The purpose of this study is to determine the activity of the RVG neurotoxin-like motif on nAChR subtypes that are expressed in brain regions involved in rabid animal behavior. nAChRs were expressed in Xenopus laevis oocytes, and two-electrode voltage clamp electrophysiology was used to collect concentration-response data to measure the functional effects. The RVG peptide preferentially and completely inhibits α7 nAChR ACh-induced currents by a competitive antagonist mechanism. Tested heteromeric nAChRs are also inhibited, but to a lesser extent than the α7 subtype. Residues of the RVG peptide with high sequence homology to αBTX and other neurotoxins were substituted with alanine. Altered RVG neurotoxin-like peptides showed that residues phenylalanine 192, arginine 196, and arginine 199 are important determinants of RVG peptide apparent potency on α7 nAChRs, while serine 195 is not. The evaluation of the rabies ectodomain reaffirmed the observations made with the RVG peptide, illustrating a significant inhibitory impact on α7 nAChR with potency in the nanomolar range. In a mammalian cell culture model of neurons, we confirm that the RVG peptide binds preferentially to cells expressing the α7 nAChR. Defining the activity of the RVG peptide on nAChRs expands our understanding of basic mechanisms in host-pathogen interactions that result in neurological disorders.
Keywords: N2a; RVG; alpha7; electrophysiology; nAChR; nicotinic acetylcholine receptor; rabies virus; rabies virus glycoprotein.
Copyright © 2024 O’Brien, Thao, Weber, Danielson, Boldt, Hueffer and Weltzin.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
Recombinant Newcastle disease virus rL-RVG enhances the apoptosis and inhibits the migration of A549 lung adenocarcinoma cells via regulating alpha 7 nicotinic acetylcholine receptors in vitro.Virol J. 2017 Oct 3;14(1):190. doi: 10.1186/s12985-017-0852-z. Virol J. 2017. PMID: 28974241 Free PMC article.
-
Migration of gastric cancer is suppressed by recombinant Newcastle disease virus (rL-RVG) via regulating α7-nicotinic acetylcholine receptors/ERK- EMT.BMC Cancer. 2019 Oct 22;19(1):976. doi: 10.1186/s12885-019-6225-9. BMC Cancer. 2019. PMID: 31640627 Free PMC article.
-
SARS-CoV-2 spike ectodomain targets α7 nicotinic acetylcholine receptors.J Biol Chem. 2023 May;299(5):104707. doi: 10.1016/j.jbc.2023.104707. Epub 2023 Apr 13. J Biol Chem. 2023. PMID: 37061001 Free PMC article.
-
Synthetic peptides in the study of the interaction of rabies virus and the acetylcholine receptor.Adv Biochem Psychopharmacol. 1988;44:57-71. Adv Biochem Psychopharmacol. 1988. PMID: 3041753 Review.
-
The potential use of rabies virus glycoprotein-derived peptides to facilitate drug delivery into the central nervous system: a mini review.J Drug Target. 2017 Jun;25(5):379-385. doi: 10.1080/1061186X.2016.1223676. Epub 2016 Aug 31. J Drug Target. 2017. PMID: 27581650 Review.
References
-
- Antil-Delbeke S., Gaillard C., Tamiya T., Corringer P. J., Changeux J. P., Servent D., et al. . (2000). Molecular determinants by which a long chain toxin from snake venom interacts with the neuronal alpha 7-nicotinic acetylcholine receptor. J. Biol. Chem. 275, 29594–29601. doi: 10.1074/jbc.M909746199 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases