

Legionella effectors SidC/SdcA ubiquitinate multiple small GTPases and SNARE proteins to promote phagosomal maturation
- PMID: 38836877
- PMCID: PMC11335287
- DOI: 10.1007/s00018-024-05271-7
Legionella effectors SidC/SdcA ubiquitinate multiple small GTPases and SNARE proteins to promote phagosomal maturation
Abstract
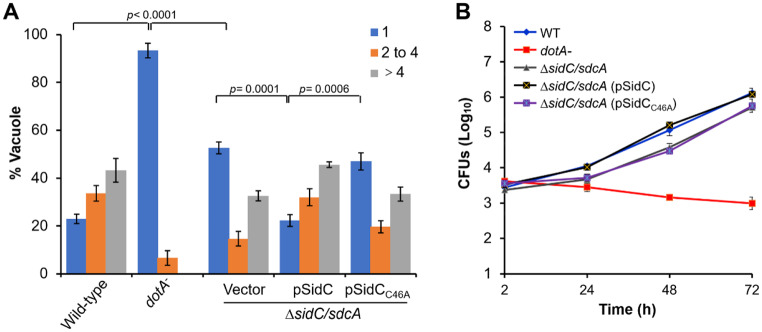
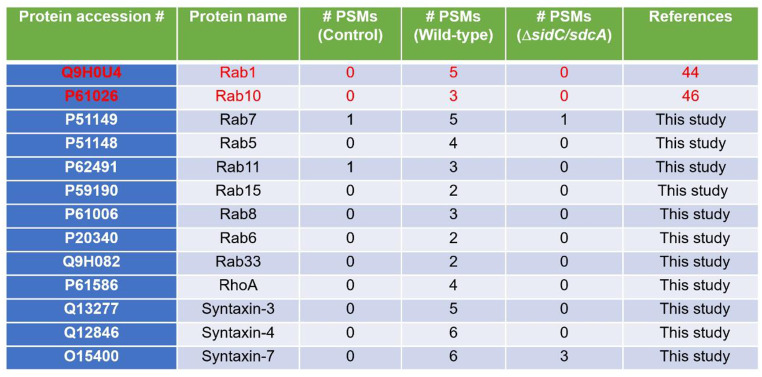
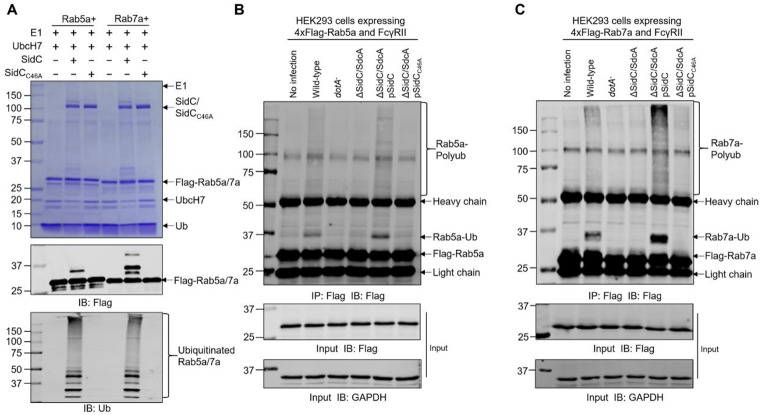
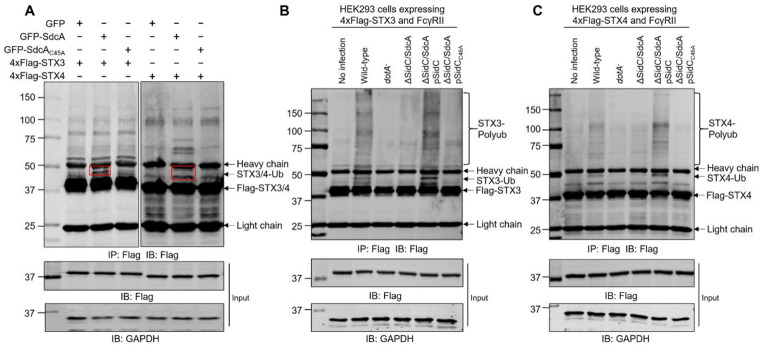
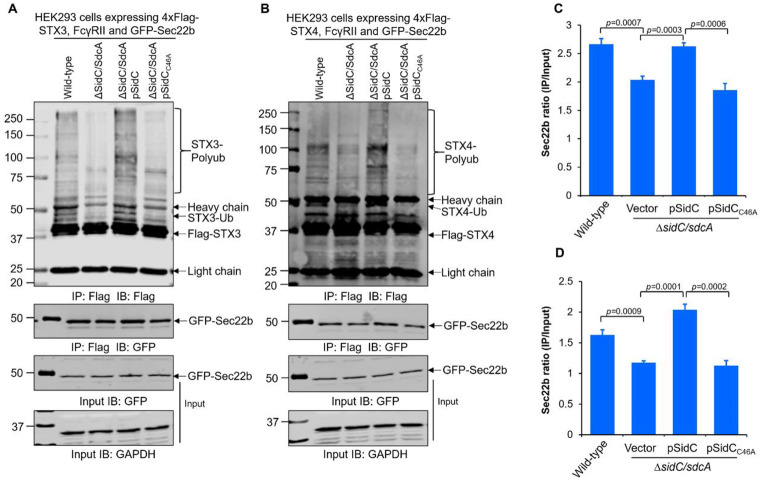
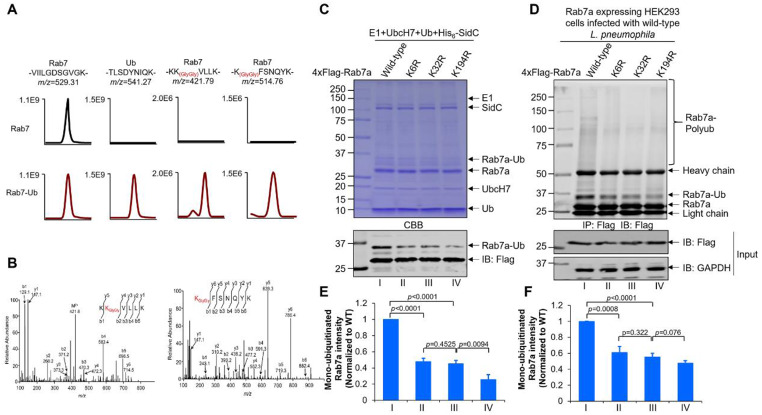
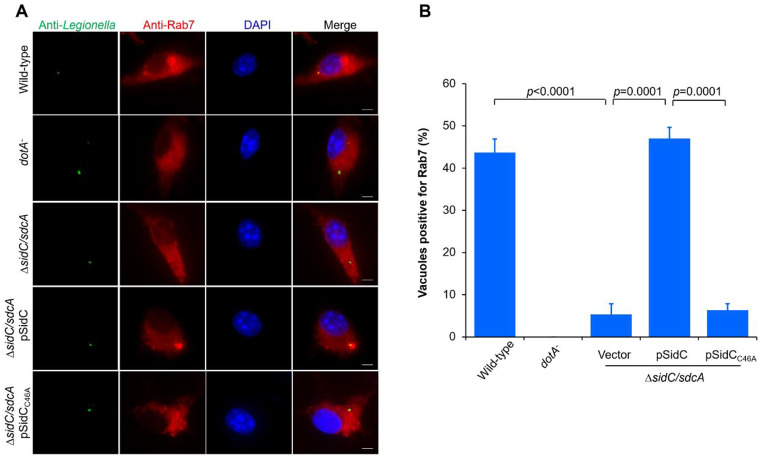
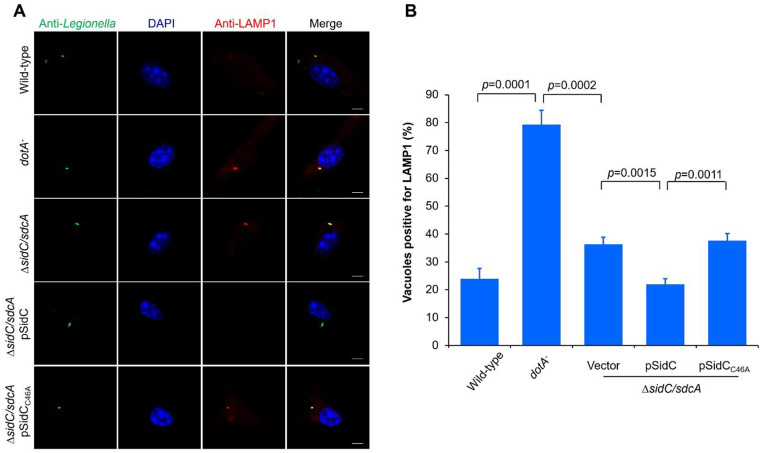
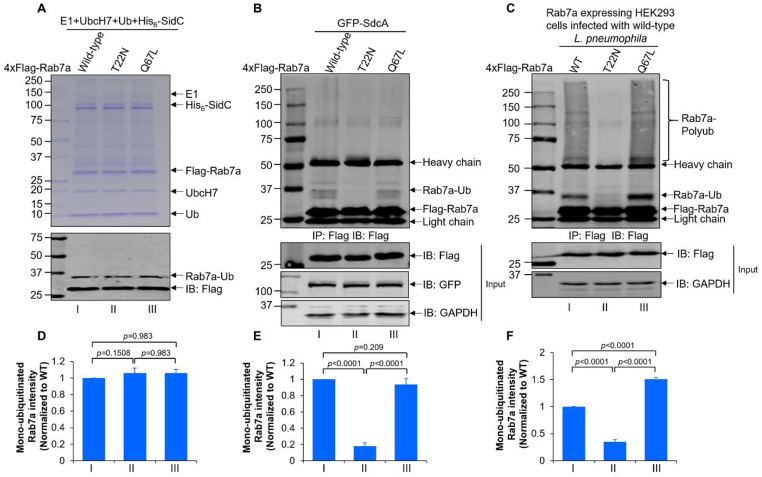
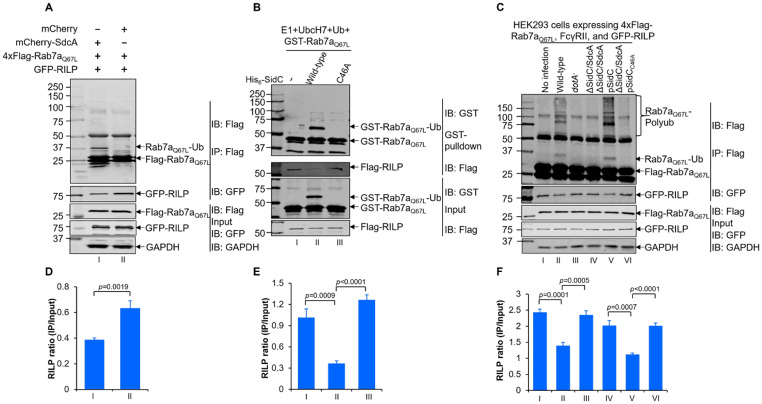
Protein ubiquitination is one of the most important posttranslational modifications (PTMs) in eukaryotes and is involved in the regulation of almost all cellular signaling pathways. The intracellular bacterial pathogen Legionella pneumophila translocates at least 26 effectors to hijack host ubiquitination signaling via distinct mechanisms. Among these effectors, SidC/SdcA are novel E3 ubiquitin ligases with the adoption of a Cys-His-Asp catalytic triad. SidC/SdcA are critical for the recruitment of endoplasmic reticulum (ER)-derived vesicles to the Legionella-containing vacuole (LCV). However, the ubiquitination targets of SidC/SdcA are largely unknown, which restricts our understanding of the mechanisms used by these effectors to hijack the vesicle trafficking pathway. Here, we demonstrated that multiple Rab small GTPases and target soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNARE) proteins are bona fide ubiquitination substrates of SidC/SdcA. SidC/SdcA-mediated ubiquitination of syntaxin 3 and syntaxin 4 promotes their unconventional pairing with the vesicle-SNARE protein Sec22b, thereby contributing to the membrane fusion of ER-derived vesicles with the phagosome. In addition, our data reveal that ubiquitination of Rab7 by SidC/SdcA is critical for its association with the LCV membrane. Rab7 ubiquitination could impair its binding with the downstream effector Rab-interacting lysosomal protein (RILP), which partially explains why LCVs avoid fusion with lysosomes despite the acquisition of Rab7. Taken together, our study reveals the biological mechanisms employed by SidC/SdcA to promote the maturation of the LCVs.
Keywords: Effector proteins; Legionella pneumophila; RILP; Rab small GTPase; SNARE proteins; Ubiquitination.
© 2024. The Author(s).
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
