

Gene regulatory landscape of cerebral cortex folding
- PMID: 38838158
- PMCID: PMC11152136
- DOI: 10.1126/sciadv.adn1640
Gene regulatory landscape of cerebral cortex folding
Abstract
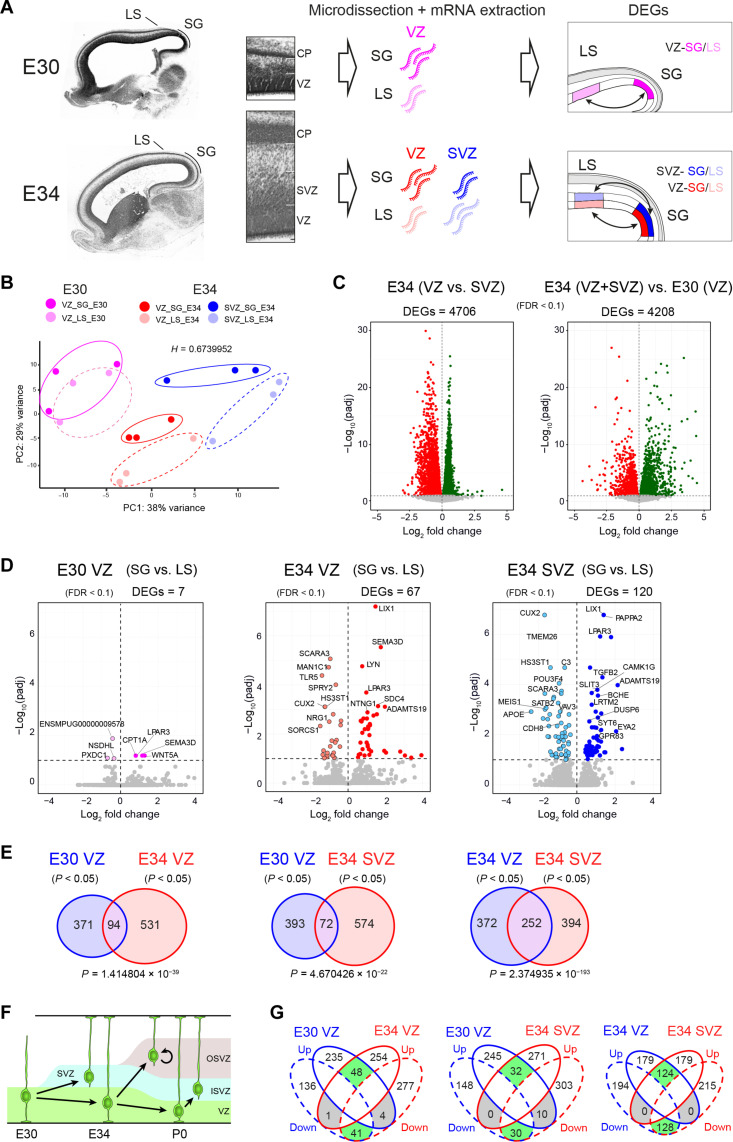
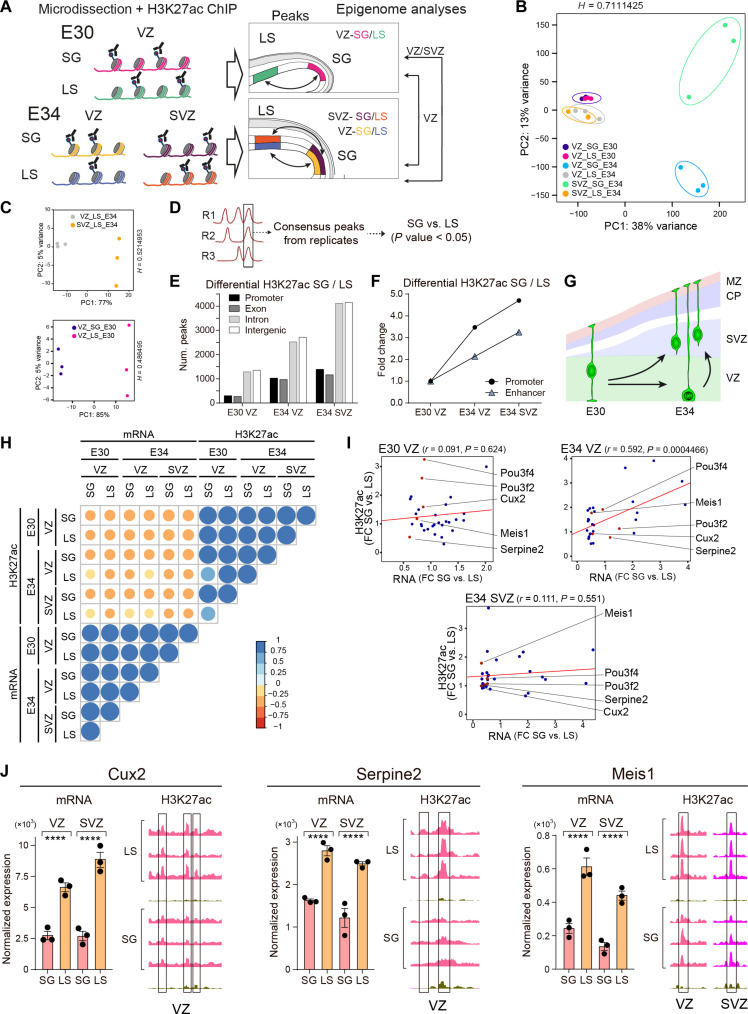
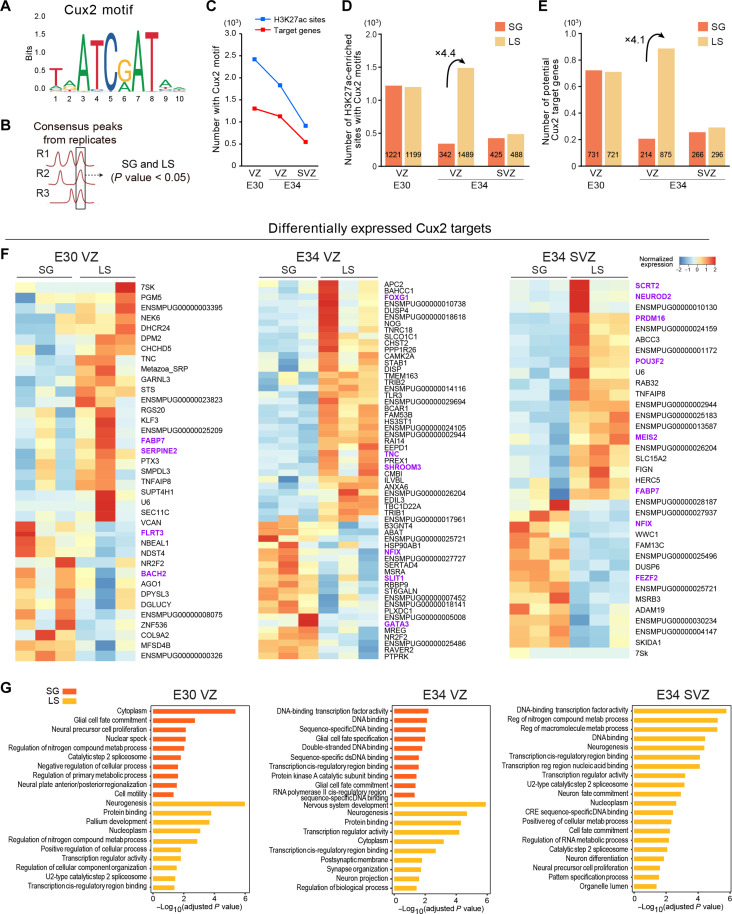
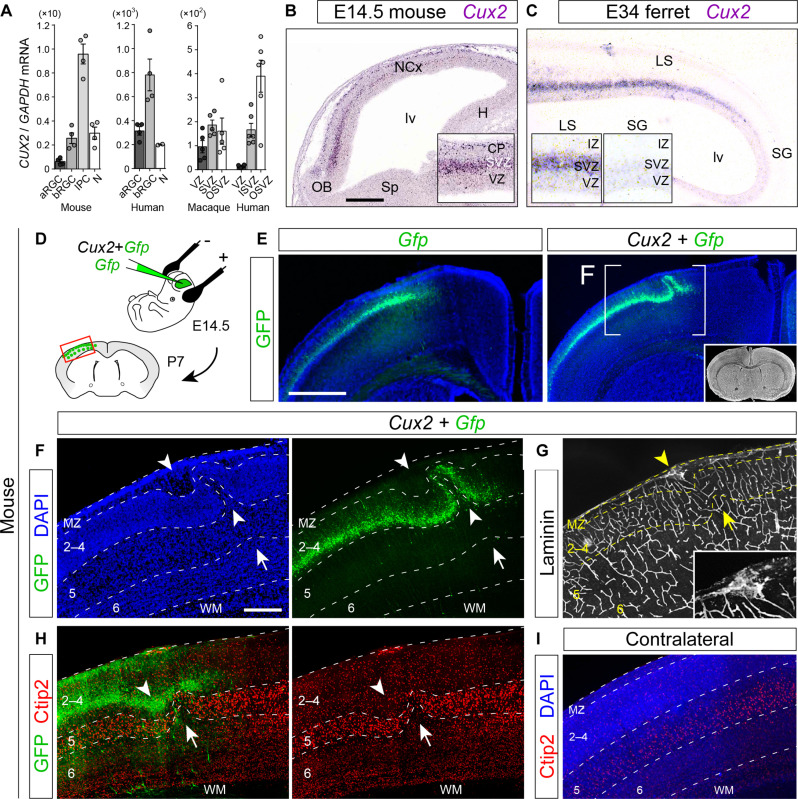
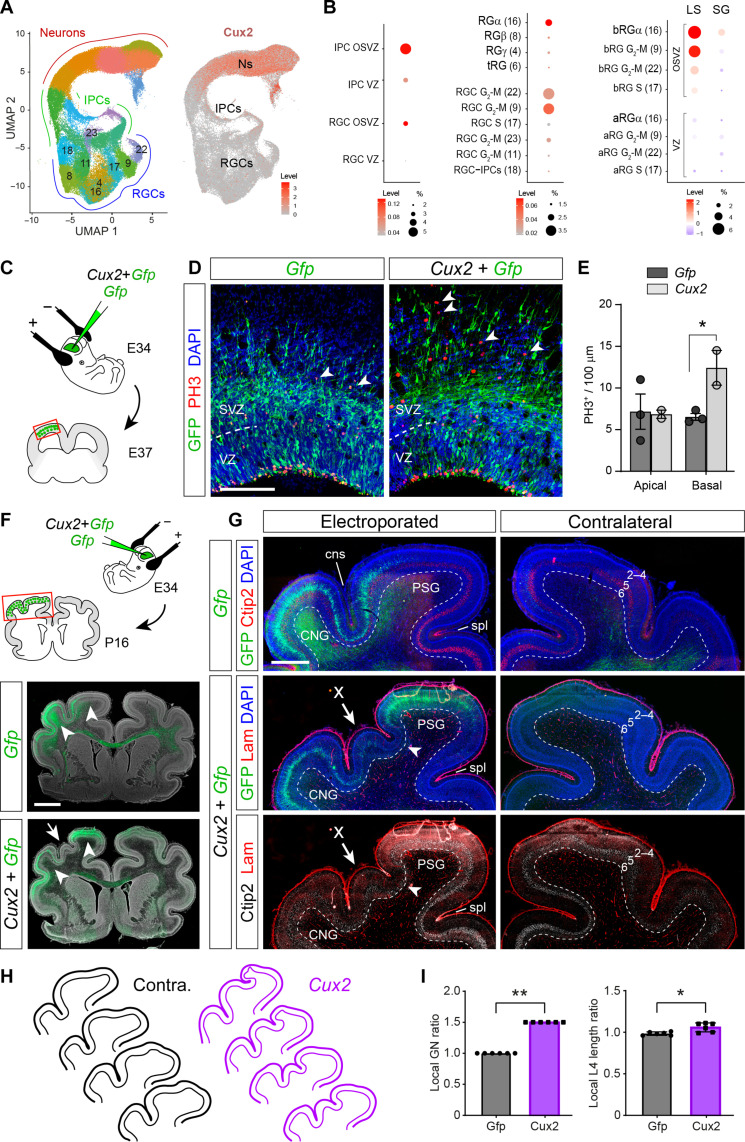
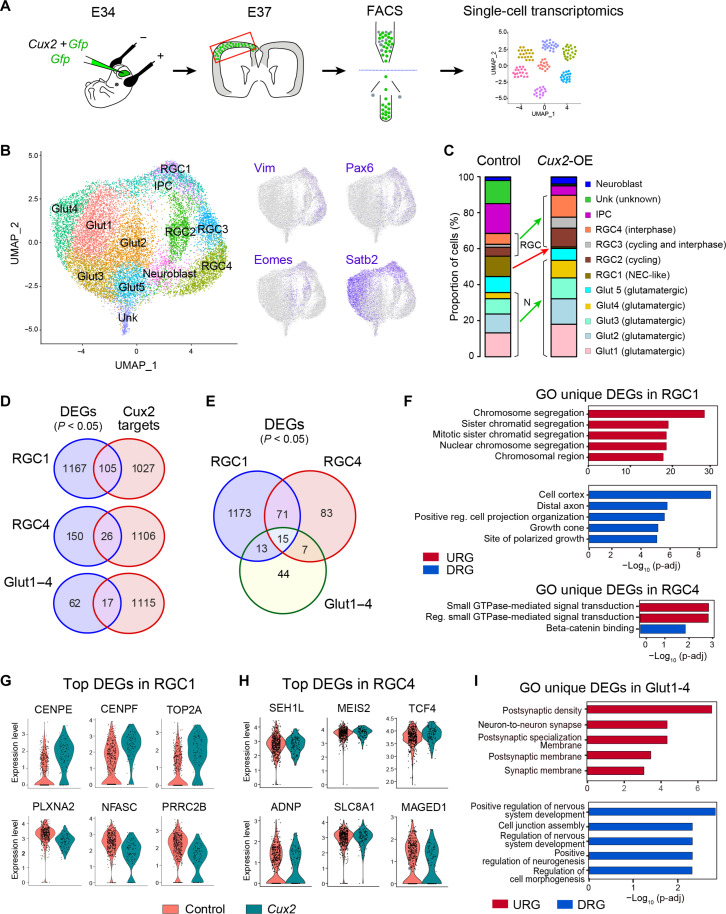
Folding of the cerebral cortex is a key aspect of mammalian brain development and evolution, and defects are linked to severe neurological disorders. Primary folding occurs in highly stereotyped patterns that are predefined in the cortical germinal zones by a transcriptomic protomap. The gene regulatory landscape governing the emergence of this folding protomap remains unknown. We characterized the spatiotemporal dynamics of gene expression and active epigenetic landscape (H3K27ac) across prospective folds and fissures in ferret. Our results show that the transcriptomic protomap begins to emerge at early embryonic stages, and it involves cell-fate signaling pathways. The H3K27ac landscape reveals developmental cell-fate restriction and engages known developmental regulators, including the transcription factor Cux2. Manipulating Cux2 expression in cortical progenitors changed their proliferation and the folding pattern in ferret, caused by selective transcriptional changes as revealed by single-cell RNA sequencing analyses. Our findings highlight the key relevance of epigenetic mechanisms in defining the patterns of cerebral cortex folding.
Figures
References
-
- W. Welker, in Cerebral Cortex, A. Peters, E. G. Jones, Eds. (Plenum Press, 1990), vol. 8B, pp. 3–136.
-
- Llinares-Benadero C., Borrell V., Deconstructing cortical folding: Genetic, cellular and mechanical determinants. Nat. Rev. Neurosci. 20, 161–176 (2019). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
