

Interferon-γ induces combined pyroptotic angiopathy and APOL1 expression in human kidney disease
- PMID: 38838223
- PMCID: PMC11216883
- DOI: 10.1016/j.celrep.2024.114310
Interferon-γ induces combined pyroptotic angiopathy and APOL1 expression in human kidney disease
Abstract
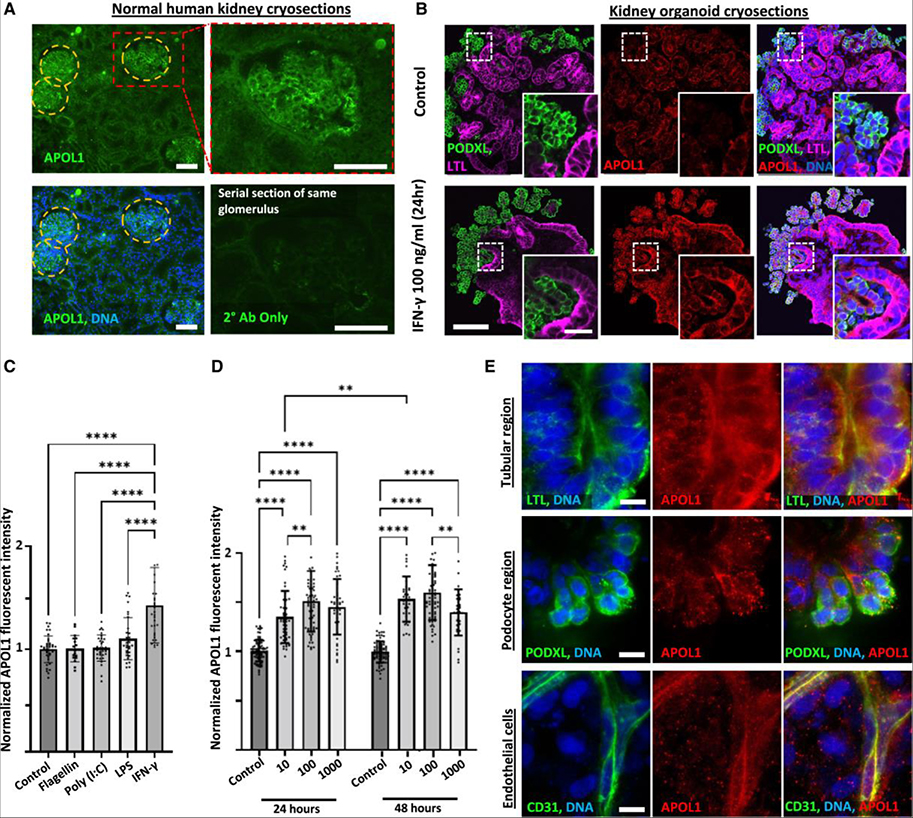
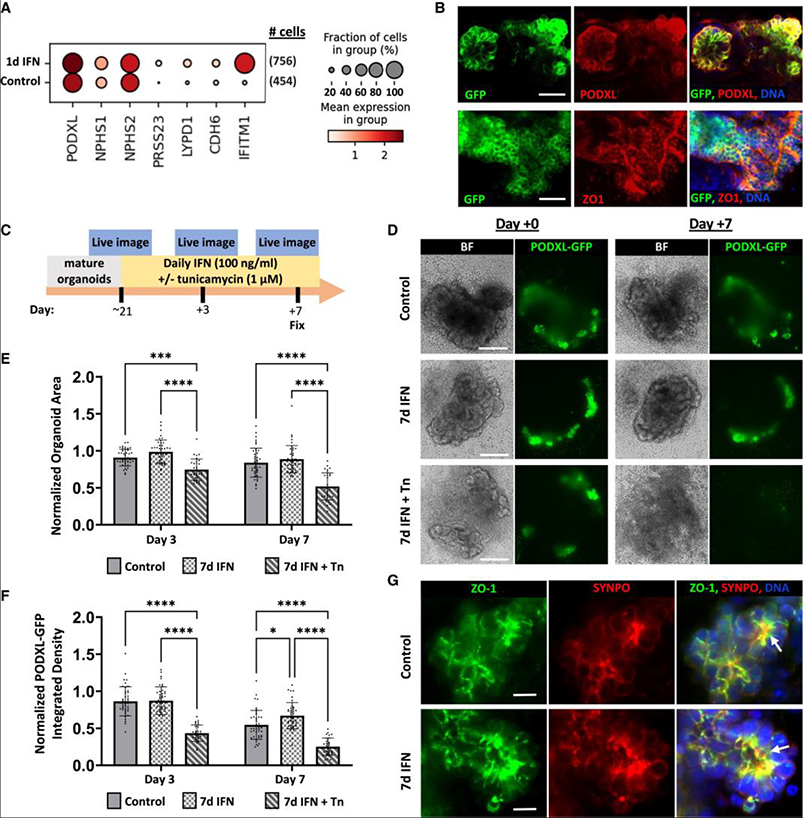
Elevated interferon (IFN) signaling is associated with kidney diseases including COVID-19, HIV, and apolipoprotein-L1 (APOL1) nephropathy, but whether IFNs directly contribute to nephrotoxicity remains unclear. Using human kidney organoids, primary endothelial cells, and patient samples, we demonstrate that IFN-γ induces pyroptotic angiopathy in combination with APOL1 expression. Single-cell RNA sequencing, immunoblotting, and quantitative fluorescence-based assays reveal that IFN-γ-mediated expression of APOL1 is accompanied by pyroptotic endothelial network degradation in organoids. Pharmacological blockade of IFN-γ signaling inhibits APOL1 expression, prevents upregulation of pyroptosis-associated genes, and rescues vascular networks. Multiomic analyses in patients with COVID-19, proteinuric kidney disease, and collapsing glomerulopathy similarly demonstrate increased IFN signaling and pyroptosis-associated gene expression correlating with accelerated renal disease progression. Our results reveal that IFN-γ signaling simultaneously induces endothelial injury and primes renal cells for pyroptosis, suggesting a combinatorial mechanism for APOL1-mediated collapsing glomerulopathy, which can be targeted therapeutically.
Keywords: CP: Cell biology; CP: Immunology; baricitinib; caspase; cell death; gasdermin; glomerulosclerosis; inflammation; podocalyxin; proteomics; pyroptosis; spatial transcriptomics.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.S.F. is an inventor on patents and/or patent applications related to human kidney organoid differentiation and modeling of disease in this system (these include “Three-dimensional differentiation of epiblast spheroids into kidney tubular organoids modeling human microphysiology, toxicology, and morphogenesis” [Japan, US, and Australia], licensed to STEMCELL Technologies; “High-throughput automation of organoids for identifying therapeutic strategies” [PTC patent application pending]; and “Systems and methods for characterizing pathophysiology” [PTC patent application pending]). B.S.F. and H.F. hold ownership interest in Plurexa, LLC. None of the preceding interests affected in any way the results of the paper or would be affected by them but are shared by way of transparency.
Figures





References
-
- Bonventre JV, and Zuk A (2004). Ischemic acute renal failure: an inflammatory disease? Kidney Int. 66, 480–485. - PubMed
-
- Evans RA, Leavy OC, Richardson M, Elneima O, McAuley HJC, Shikotra A, Singapuri A, Sereno M, Saunders RM, Harris VC, et al. (2022). Clinical characteristics with inflammation profiling of long COVID and association with 1-year recovery following hospitalisation in the UK: a prospective observational study. Lancet Respir. Med. 10, 761–775. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UH3 TR000504/TR/NCATS NIH HHS/United States
- UC2 DK126006/DK/NIDDK NIH HHS/United States
- UH3 TR002158/TR/NCATS NIH HHS/United States
- UH3 TR003288/TR/NCATS NIH HHS/United States
- R35 GM149516/GM/NIGMS NIH HHS/United States
- U2C TR004867/TR/NCATS NIH HHS/United States
- U01 AI176460/AI/NIAID NIH HHS/United States
- R01 HL167688/HL/NHLBI NIH HHS/United States
- R01 DK117914/DK/NIDDK NIH HHS/United States
- R01 DK130386/DK/NIDDK NIH HHS/United States
- TL1 DK132769/DK/NIDDK NIH HHS/United States
- U01 DK127553/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
