

Cell-class-specific electric field entrainment of neural activity
- PMID: 38838670
- PMCID: PMC11309920
- DOI: 10.1016/j.neuron.2024.05.009
Cell-class-specific electric field entrainment of neural activity
Erratum in
-
Cell-class-specific electric field entrainment of neural activity.Neuron. 2024 Aug 7;112(15):2659-2660. doi: 10.1016/j.neuron.2024.07.011. Epub 2024 Jul 26. Neuron. 2024. PMID: 39067447 Free PMC article. No abstract available.
Abstract
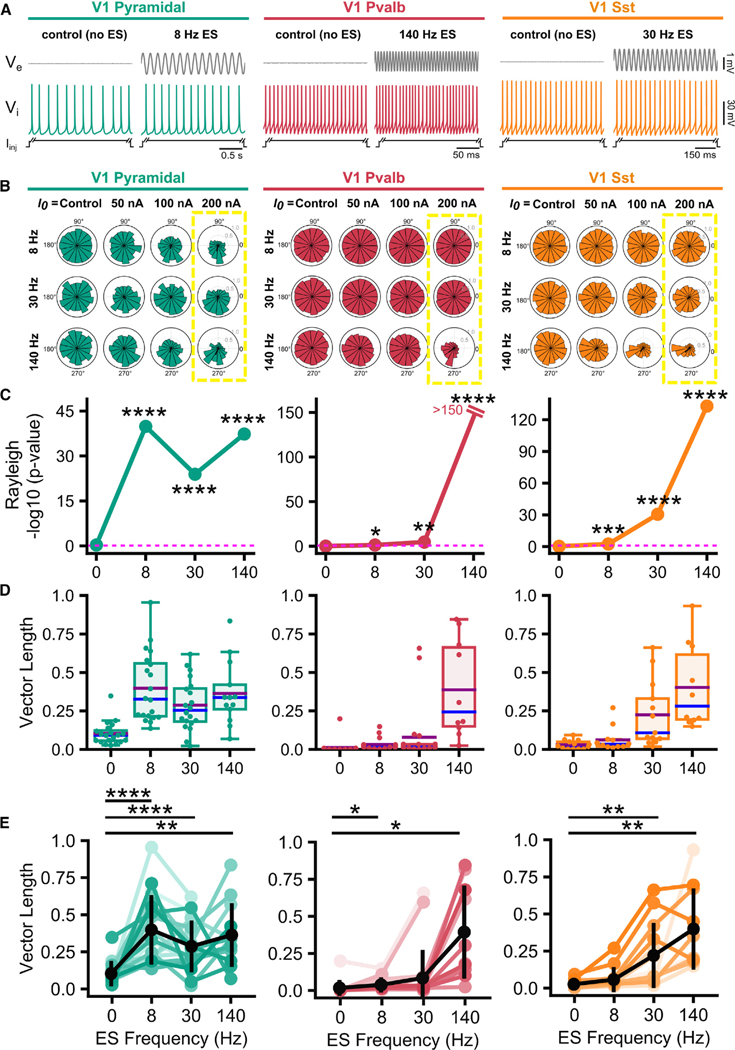
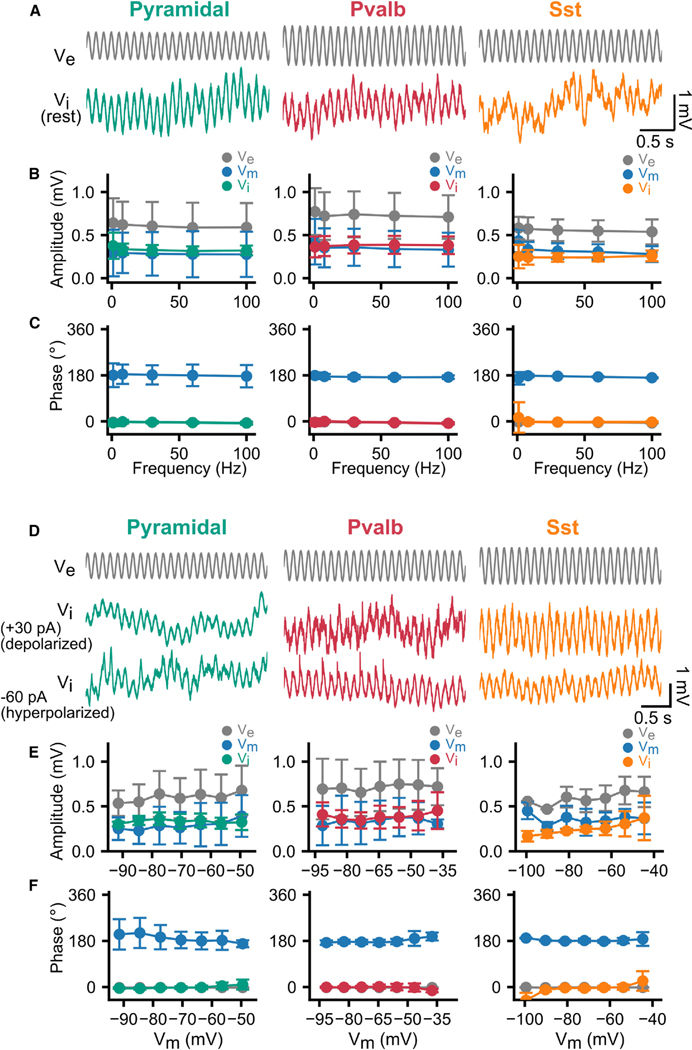
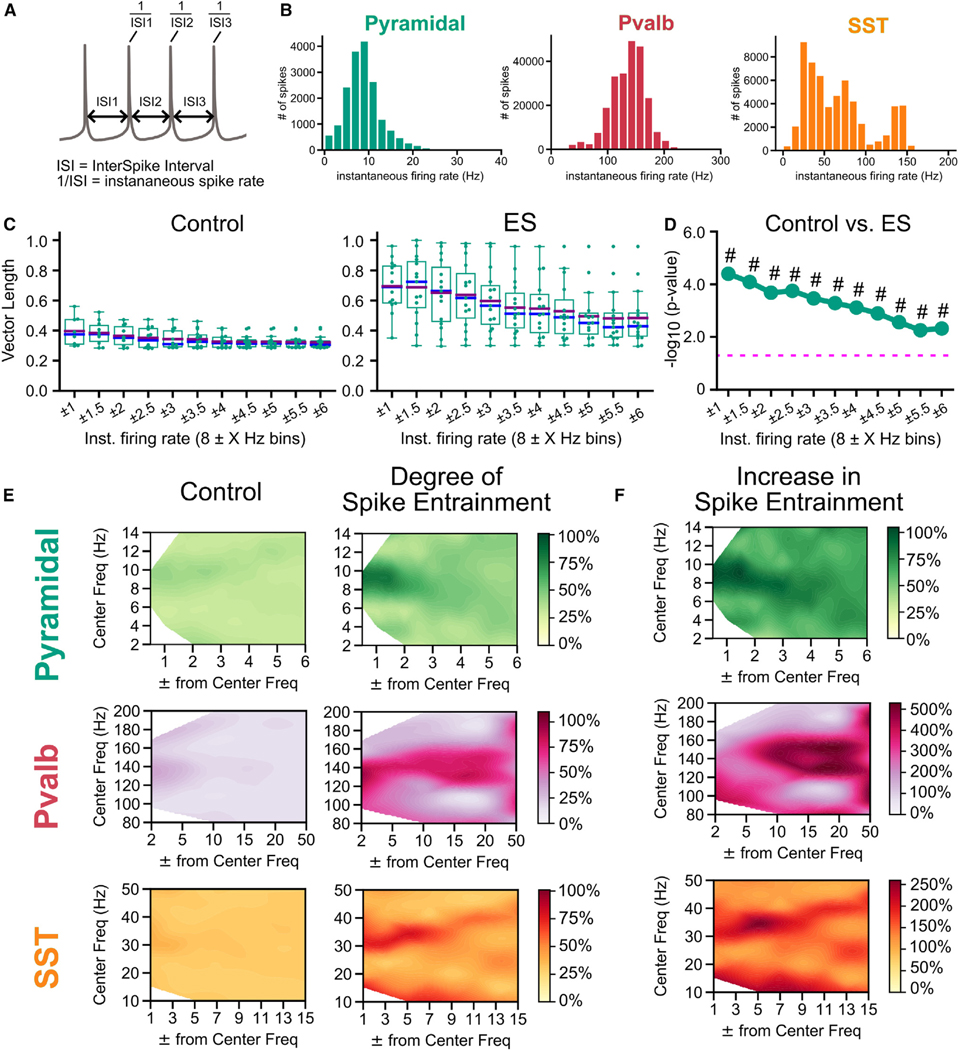
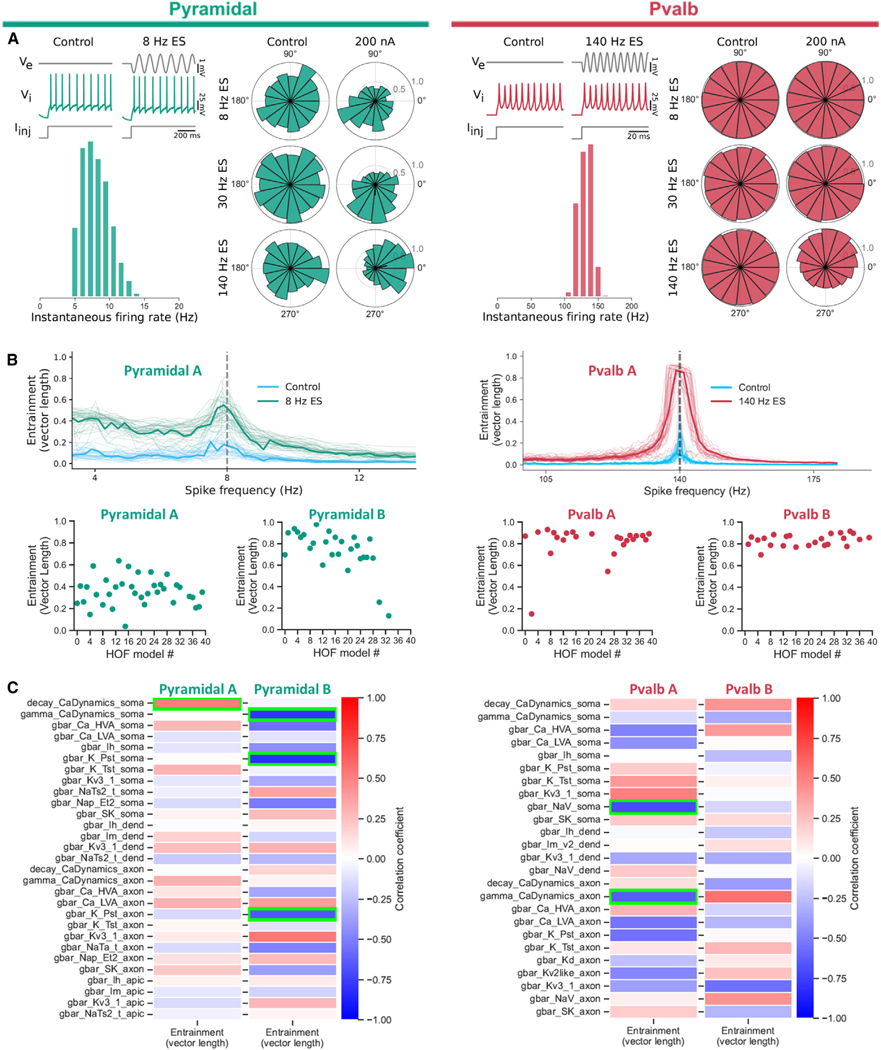
Electric fields affect the activity of neurons and brain circuits, yet how this happens at the cellular level remains enigmatic. Lack of understanding of how to stimulate the brain to promote or suppress specific activity significantly limits basic research and clinical applications. Here, we study how electric fields impact subthreshold and spiking properties of major cortical neuronal classes. We find that neurons in the rodent and human cortex exhibit strong, cell-class-dependent entrainment that depends on stimulation frequency. Excitatory pyramidal neurons, with their slower spike rate, entrain to both slow and fast electric fields, while inhibitory classes like Pvalb and Sst (with their fast spiking) predominantly phase-lock to fast fields. We show that this spike-field entrainment is the result of two effects: non-specific membrane polarization occurring across classes and class-specific excitability properties. Importantly, these properties are present across cortical areas and species. These findings allow for the design of selective and class-specific neuromodulation.
Keywords: behavior; cell types; characterization; cre-animals; electric field; entrainment; human; mouse; patch-clamp; simulation; stimulation.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.Y.L. has previously consulted for Starfish Neuroscience, Inc. C.A.A. and S.Y.L. are listed as inventors on a patent application related to this work. C.K. is a board member of and has a financial interest in Intrinsic Powers, Inc.
Figures


Update of
-
Cell class-specific electric field entrainment of neural activity.bioRxiv [Preprint]. 2024 Mar 6:2023.02.14.528526. doi: 10.1101/2023.02.14.528526. bioRxiv. 2024. Update in: Neuron. 2024 Aug 7;112(15):2614-2630.e5. doi: 10.1016/j.neuron.2024.05.009. PMID: 36824721 Free PMC article. Updated. Preprint.
References
-
- Hitzig E. (1874). Untersuchungen über das Gehirn: Abhandlungen physiologischen und pathologischen Inhalts (A. Hirschwald; ).
-
- Fritsch G. (1870). Uber die elektrische Erregbarkeit des Grosshirns. Arch. Anat. Physiol. Wiss. Med 37, 300–332.
-
- Vitek JL, Jain R, Chen L, Tröster AI, Schrock LE, House PA, Giroux ML, Hebb AO, Farris SM, Whiting DM, et al. (2020). Subthalamic nucleus deep brain stimulation with a multiple independent constant current-controlled device in Parkinson’s disease (INTREPID): a multicentre, double-blind, randomised, sham-controlled study. Lancet Neurol. 19, 491–501. 10.1016/S1474-4422(20)30108-3. - DOI - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
