

Beyond homogeneity: Assessing the validity of the Michaelis-Menten rate law in spatially heterogeneous environments
- PMID: 38843305
- PMCID: PMC11185478
- DOI: 10.1371/journal.pcbi.1012205
Beyond homogeneity: Assessing the validity of the Michaelis-Menten rate law in spatially heterogeneous environments
Abstract
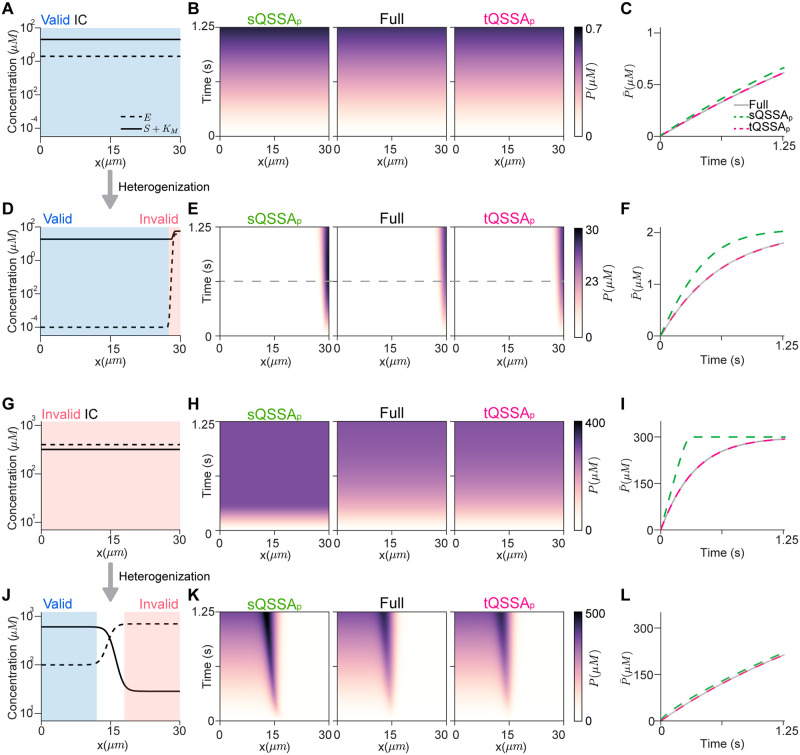
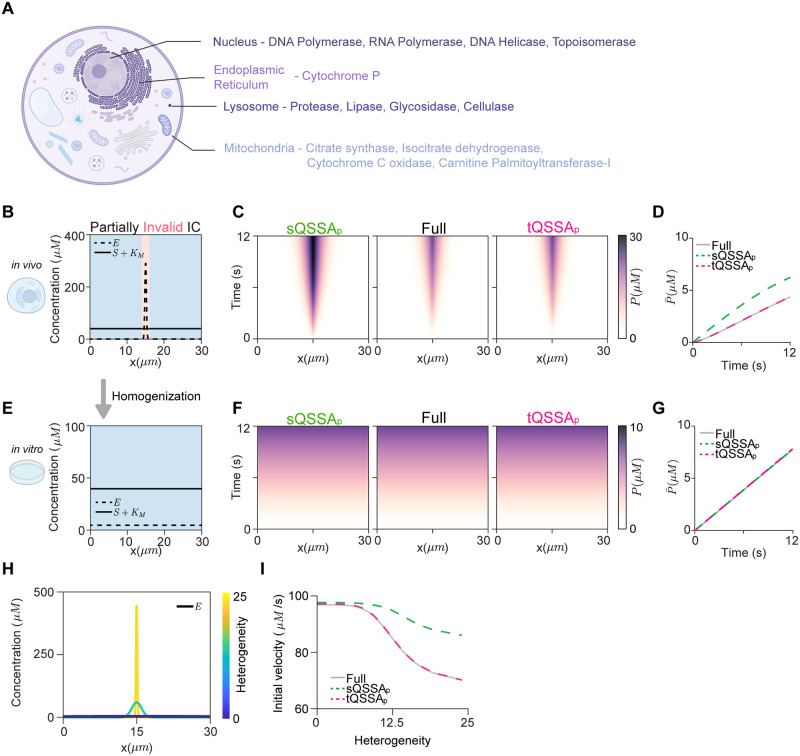
The Michaelis-Menten (MM) rate law has been a fundamental tool in describing enzyme-catalyzed reactions for over a century. When substrates and enzymes are homogeneously distributed, the validity of the MM rate law can be easily assessed based on relative concentrations: the substrate is in large excess over the enzyme-substrate complex. However, the applicability of this conventional criterion remains unclear when species exhibit spatial heterogeneity, a prevailing scenario in biological systems. Here, we explore the MM rate law's applicability under spatial heterogeneity by using partial differential equations. In this study, molecules diffuse very slowly, allowing them to locally reach quasi-steady states. We find that the conventional criterion for the validity of the MM rate law cannot be readily extended to heterogeneous environments solely through spatial averages of molecular concentrations. That is, even when the conventional criterion for the spatial averages is satisfied, the MM rate law fails to capture the enzyme catalytic rate under spatial heterogeneity. In contrast, a slightly modified form of the MM rate law, based on the total quasi-steady state approximation (tQSSA), is accurate. Specifically, the tQSSA-based modified form, but not the original MM rate law, accurately predicts the drug clearance via cytochrome P450 enzymes and the ultrasensitive phosphorylation in heterogeneous environments. Our findings shed light on how to simplify spatiotemporal models for enzyme-catalyzed reactions in the right context, ensuring accurate conclusions and avoiding misinterpretations in in silico simulations.
Copyright: © 2024 Shin et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Misuse of the Michaelis-Menten rate law for protein interaction networks and its remedy.PLoS Comput Biol. 2020 Oct 22;16(10):e1008258. doi: 10.1371/journal.pcbi.1008258. eCollection 2020 Oct. PLoS Comput Biol. 2020. PMID: 33090989 Free PMC article. Review.
-
Quasi-steady-state kinetics at enzyme and substrate concentrations in excess of the Michaelis-Menten constant.J Theor Biol. 2007 Apr 21;245(4):737-48. doi: 10.1016/j.jtbi.2006.12.005. Epub 2006 Dec 12. J Theor Biol. 2007. PMID: 17234216
-
Michaelis-Menten kinetics at high enzyme concentrations.Bull Math Biol. 2003 Nov;65(6):1111-29. doi: 10.1016/S0092-8240(03)00059-4. Bull Math Biol. 2003. PMID: 14607291
-
The total quasi-steady-state approximation is valid for reversible enzyme kinetics.J Theor Biol. 2004 Feb 7;226(3):303-13. doi: 10.1016/j.jtbi.2003.09.006. J Theor Biol. 2004. PMID: 14643644
-
Validity of the Michaelis-Menten equation--steady-state or reactant stationary assumption: that is the question.FEBS J. 2014 Jan;281(2):464-72. doi: 10.1111/febs.12564. Epub 2013 Nov 18. FEBS J. 2014. PMID: 24245583 Review.
Cited by
-
From homogeneity to heterogeneity: Refining stochastic simulations of gene regulation.Comput Struct Biotechnol J. 2025 Jan 15;27:411-422. doi: 10.1016/j.csbj.2025.01.004. eCollection 2025. Comput Struct Biotechnol J. 2025. PMID: 39906159 Free PMC article.
-
Optimizing enzyme inhibition analysis: precise estimation with a single inhibitor concentration.Nat Commun. 2025 Jun 5;16(1):5217. doi: 10.1038/s41467-025-60468-z. Nat Commun. 2025. PMID: 40473596 Free PMC article.
References
-
- Griffiths AJ. Modern genetic analysis: integrating genes and genomes. vol. 1. Macmillan; 2002.
-
- Cooper GM, Adams KW. The cell: a molecular approach. Oxford University Press; 2023.
-
- Henri V. Lois générales de l’action des diastases. Librairie Scientifique A. Hermann; 1903.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
