

Development and organization of the retinal orientation selectivity map
- PMID: 38844438
- PMCID: PMC11156980
- DOI: 10.1038/s41467-024-49206-z
Development and organization of the retinal orientation selectivity map
Abstract
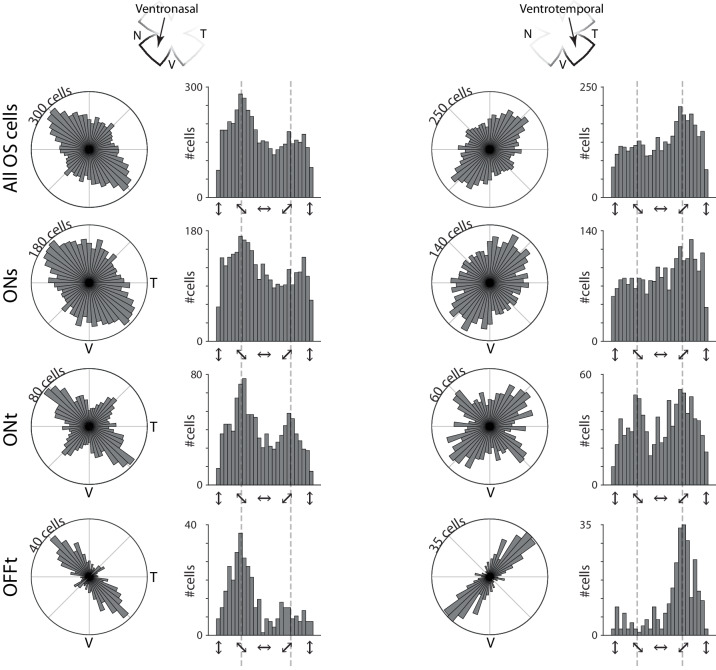
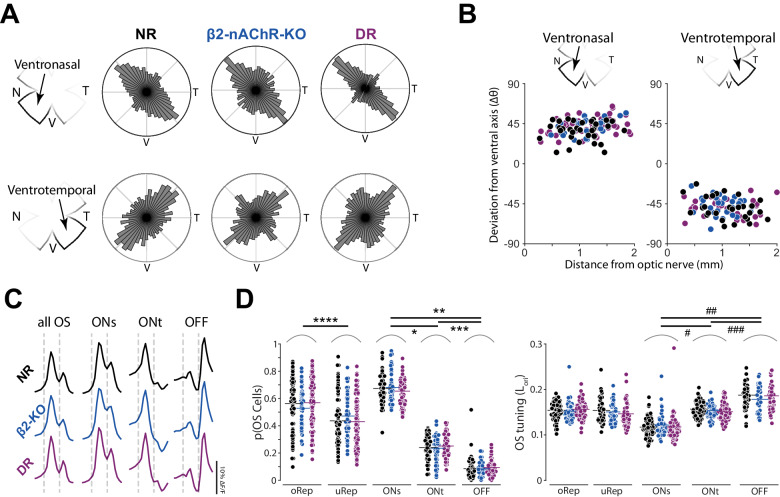
Orientation or axial selectivity, the property of neurons in the visual system to respond preferentially to certain angles of visual stimuli, plays a pivotal role in our understanding of visual perception and information processing. This computation is performed as early as the retina, and although much work has established the cellular mechanisms of retinal orientation selectivity, how this computation is organized across the retina is unknown. Using a large dataset collected across the mouse retina, we demonstrate functional organization rules of retinal orientation selectivity. First, we identify three major functional classes of retinal cells that are orientation selective and match previous descriptions. Second, we show that one orientation is predominantly represented in the retina and that this predominant orientation changes as a function of retinal location. Third, we demonstrate that neural activity plays little role on the organization of retinal orientation selectivity. Lastly, we use in silico modeling followed by validation experiments to demonstrate that the overrepresented orientation aligns along concentric axes. These results demonstrate that, similar to direction selectivity, orientation selectivity is organized in a functional map as early as the retina.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Update of
-
Development and Organization of the Retinal Orientation Selectivity Map.bioRxiv [Preprint]. 2024 Mar 28:2024.03.27.585774. doi: 10.1101/2024.03.27.585774. bioRxiv. 2024. Update in: Nat Commun. 2024 Jun 6;15(1):4829. doi: 10.1038/s41467-024-49206-z. PMID: 38585937 Free PMC article. Updated. Preprint.
Similar articles
-
Development and Organization of the Retinal Orientation Selectivity Map.bioRxiv [Preprint]. 2024 Mar 28:2024.03.27.585774. doi: 10.1101/2024.03.27.585774. bioRxiv. 2024. Update in: Nat Commun. 2024 Jun 6;15(1):4829. doi: 10.1038/s41467-024-49206-z. PMID: 38585937 Free PMC article. Updated. Preprint.
-
Cardinal Orientation Selectivity Is Represented by Two Distinct Ganglion Cell Types in Mouse Retina.J Neurosci. 2016 Mar 16;36(11):3208-21. doi: 10.1523/JNEUROSCI.4554-15.2016. J Neurosci. 2016. PMID: 26985031 Free PMC article.
-
A spherical code of retinal orientation selectivity enables decoding in ensembled and retinotopic operation.Cell Rep. 2025 Mar 25;44(3):115373. doi: 10.1016/j.celrep.2025.115373. Epub 2025 Mar 1. Cell Rep. 2025. PMID: 40023844
-
Direction selectivity in the retina: symmetry and asymmetry in structure and function.Nat Rev Neurosci. 2012 Feb 8;13(3):194-208. doi: 10.1038/nrn3165. Nat Rev Neurosci. 2012. PMID: 22314444 Review.
-
Stimulus-dependent engagement of neural mechanisms for reliable motion detection in the mouse retina.J Neurophysiol. 2018 Sep 1;120(3):1153-1161. doi: 10.1152/jn.00716.2017. Epub 2018 Jun 13. J Neurophysiol. 2018. PMID: 29897862 Free PMC article. Review.
Cited by
-
Experience influences the refinement of feature selectivity in the mouse primary visual thalamus.Neuron. 2025 May 7;113(9):1352-1362.e4. doi: 10.1016/j.neuron.2025.02.023. Epub 2025 Mar 19. Neuron. 2025. PMID: 40112812
-
Compartmentalized pooling generates orientation selectivity in wide-field amacrine cells.Proc Natl Acad Sci U S A. 2024 Dec 3;121(49):e2411130121. doi: 10.1073/pnas.2411130121. Epub 2024 Nov 27. Proc Natl Acad Sci U S A. 2024. PMID: 39602271 Free PMC article.
-
Orientation selectivity properties for the affine Gaussian derivative and the affine Gabor models for visual receptive fields.J Comput Neurosci. 2025 Mar;53(1):61-98. doi: 10.1007/s10827-024-00888-w. Epub 2025 Jan 29. J Comput Neurosci. 2025. PMID: 39878929 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
