

Dominant missense variants in SREBF2 are associated with complex dermatological, neurological, and skeletal abnormalities
- PMID: 38847193
- PMCID: PMC12358889
- DOI: 10.1016/j.gim.2024.101174
Dominant missense variants in SREBF2 are associated with complex dermatological, neurological, and skeletal abnormalities
Abstract
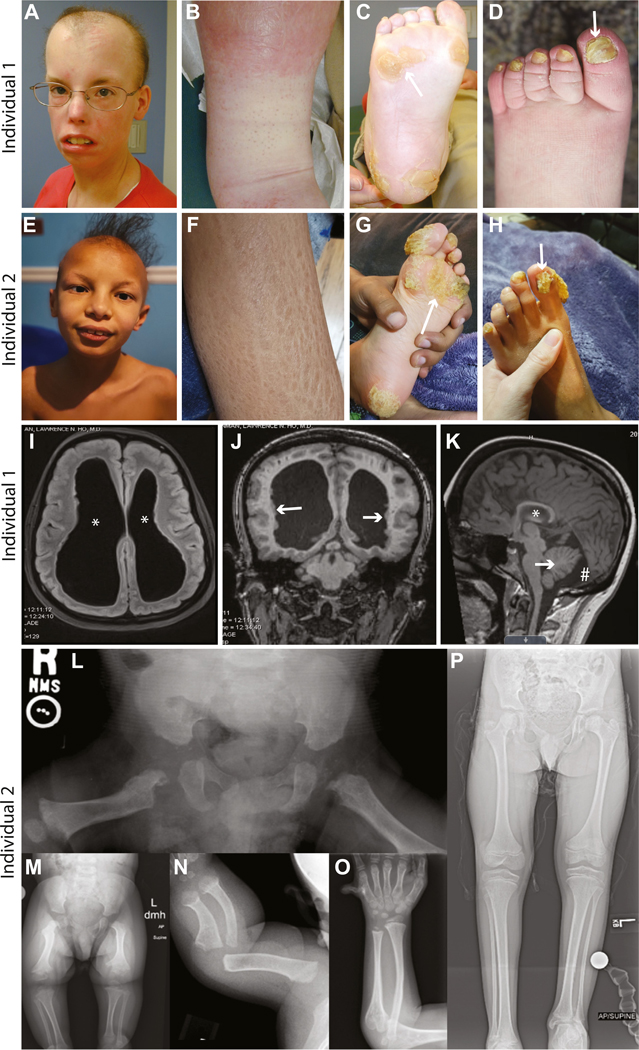
Purpose: We identified 2 individuals with de novo variants in SREBF2 that disrupt a conserved site 1 protease (S1P) cleavage motif required for processing SREBP2 into its mature transcription factor. These individuals exhibit complex phenotypic manifestations that partially overlap with sterol regulatory element binding proteins (SREBP) pathway-related disease phenotypes, but SREBF2-related disease has not been previously reported. Thus, we set out to assess the effects of SREBF2 variants on SREBP pathway activation.
Methods: We undertook ultrastructure and gene expression analyses using fibroblasts from an affected individual and utilized a fly model of lipid droplet (LD) formation to investigate the consequences of SREBF2 variants on SREBP pathway function.
Results: We observed reduced LD formation, endoplasmic reticulum expansion, accumulation of aberrant lysosomes, and deficits in SREBP2 target gene expression in fibroblasts from an affected individual, indicating that the SREBF2 variant inhibits SREBP pathway activation. Using our fly model, we discovered that SREBF2 variants fail to induce LD production and act in a dominant-negative manner, which can be rescued by overexpression of S1P.
Conclusion: Taken together, these data reveal a mechanism by which SREBF2 pathogenic variants that disrupt the S1P cleavage motif cause disease via dominant-negative antagonism of S1P, limiting the cleavage of S1P targets, including SREBP1 and SREBP2.
Keywords: Drosophila; Lipid droplet; S1P; SREBF1; SREBF2.
Copyright © 2024 American College of Medical Genetics and Genomics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of Interest The authors declare no conflicts of interest.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
