

Induction of in vivo-like ciliation in confluent monolayers of re-differentiated equine oviduct epithelial cells†
- PMID: 38847468
- PMCID: PMC11402525
- DOI: 10.1093/biolre/ioae090
Induction of in vivo-like ciliation in confluent monolayers of re-differentiated equine oviduct epithelial cells†
Abstract
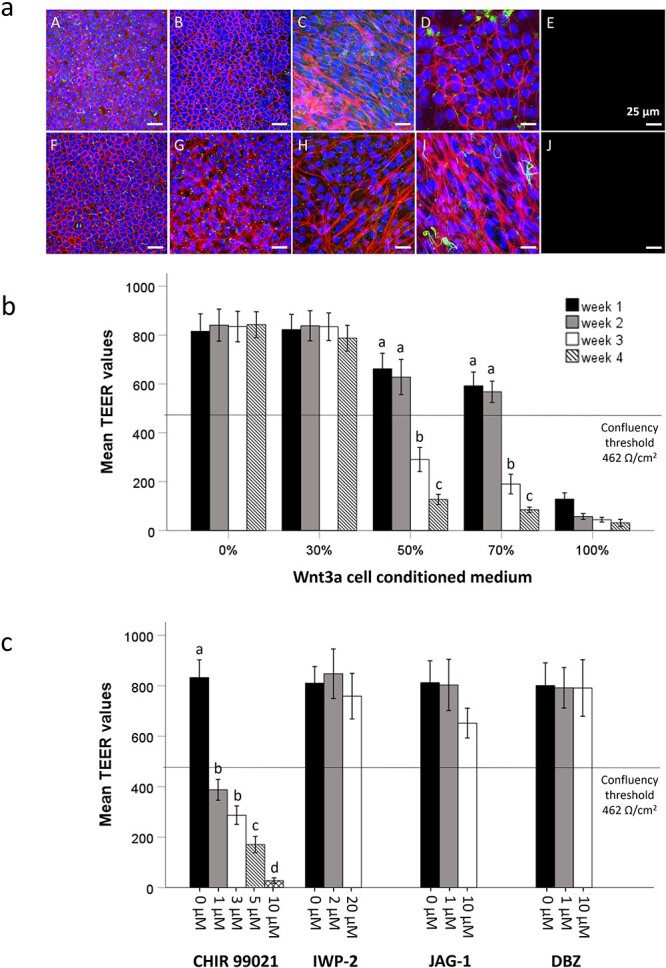
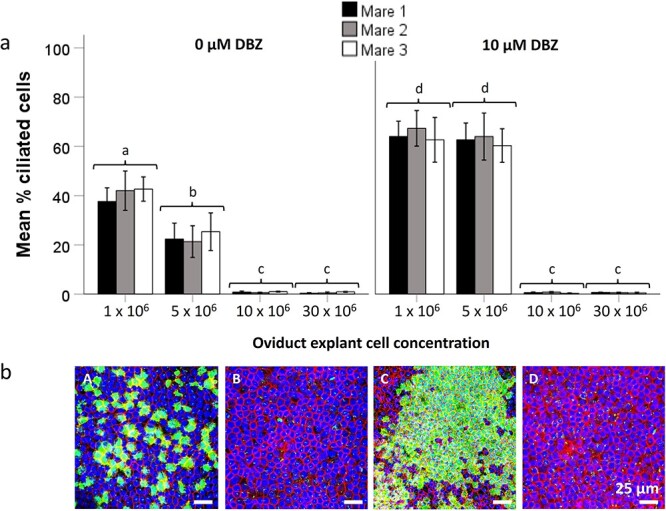
We recently developed re-differentiated equine oviduct epithelial cell (REOEC) monolayers demonstrating various in vivo morphological characteristics, but lacking secondary ciliation. In this study, we evaluated the effects of fetal bovine serum, reproductive steroid hormones, Wnt- and Notch ligands and inhibitors, and different EOEC seeding densities, in both conventional wells and on microporous membranes, on EOEC morphology and, in particular, secondary ciliation. REOEC monolayers were assessed by confocal microscopy after combined staining of nuclei, cilia, and the cytoskeleton. Only Wnt ligands, Notch inhibitors and oviduct explant cell concentration affected EOEC morphology. Undesirable epithelial-mesenchymal transition was observed in REOEC monolayers exposed to Wnt3a containing medium and Wnt ligand CHIR 99021. With respect to secondary ciliation, only the combined effect of oviduct explant cell concentration and Notch inhibition steered REOEC monolayers to in vivo-like ciliation patterns. De-differentiated EOECs, formed 10 days after oviduct explant cell seeding, were reseeded on inserts; only at initial oviduct explant cell concentrations of 1 and 5 × 106 cells per well was the formation of REOEC monolayers with a high rate of diffuse ciliation supported. Within 1 month after air-liquid interface introduction, >40% and >20% of the REOECs showed secondary cilia, respectively. At higher oviduct explant cell seeding densities secondary ciliation was not supported after re-differentiation. Additionally, Notch inhibition helped boost secondary ciliation rates to >60% in REOEC monolayers with diffuse ciliation only. These monolayers demonstrated higher clathrin expression under follicular phase conditions. Overall, the ciliated REOEC monolayers better resemble in vivo oviduct epithelial cells than previous models.
Keywords: in vitro model; ciliation; horse; oviduct.
© The Author(s) 2024. Published by Oxford University Press on behalf of Society for the Study of Reproduction.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures






Similar articles
-
Developing a reproducible protocol for culturing functional confluent monolayers of differentiated equine oviduct epithelial cells†.Biol Reprod. 2022 Apr 26;106(4):710-729. doi: 10.1093/biolre/ioab243. Biol Reprod. 2022. PMID: 34962550 Free PMC article.
-
Culture of bovine oviduct epithelial cells (BOEC).Anat Rec. 1995 Nov;243(3):347-56. doi: 10.1002/ar.1092430309. Anat Rec. 1995. PMID: 8579254
-
Long-term viability and differentiation of bovine oviductal monolayers: bidimensional versus three-dimensional culture.Theriogenology. 2012 Oct 15;78(7):1456-64. doi: 10.1016/j.theriogenology.2012.06.010. Epub 2012 Aug 24. Theriogenology. 2012. PMID: 22925649
-
In vitro systems for intercepting early embryo-maternal cross-talk in the bovine oviduct.Theriogenology. 2010 Apr 1;73(6):802-16. doi: 10.1016/j.theriogenology.2009.09.036. Epub 2009 Dec 5. Theriogenology. 2010. PMID: 19963260 Review.
-
Designing 3-Dimensional In Vitro Oviduct Culture Systems to Study Mammalian Fertilization and Embryo Production.Ann Biomed Eng. 2017 Jul;45(7):1731-1744. doi: 10.1007/s10439-016-1760-x. Epub 2016 Nov 14. Ann Biomed Eng. 2017. PMID: 27844174 Free PMC article. Review.
Cited by
-
Decoding Müllerian Duct Epithelial Regionalization.Mol Reprod Dev. 2025 Feb;92(2):e70018. doi: 10.1002/mrd.70018. Mol Reprod Dev. 2025. PMID: 39994938 Free PMC article. Review.
References
-
- Dobrinski I, Jacob JR, Tennant BC, Ball BA. Generation of an equine oviductal epithelial cell line for the study of sperm-oviduct interactions. Theriogenology 1999; 52:875–885. - PubMed
-
- Thomas PGA, Ignotz GG, Ball BA, Miller PG, Brinsko SP, Currie B. Isolation, culture, and characterization of equine oviduct epithelial-cells in-vitro. Mol Reprod Dev 1995; 41:468–478. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
