

Chaperone- and PTM-mediated activation of IRF1 tames radiation-induced cell death and the inflammatory response
- PMID: 38849539
- PMCID: PMC11291999
- DOI: 10.1038/s41423-024-01185-3
Chaperone- and PTM-mediated activation of IRF1 tames radiation-induced cell death and the inflammatory response
Abstract
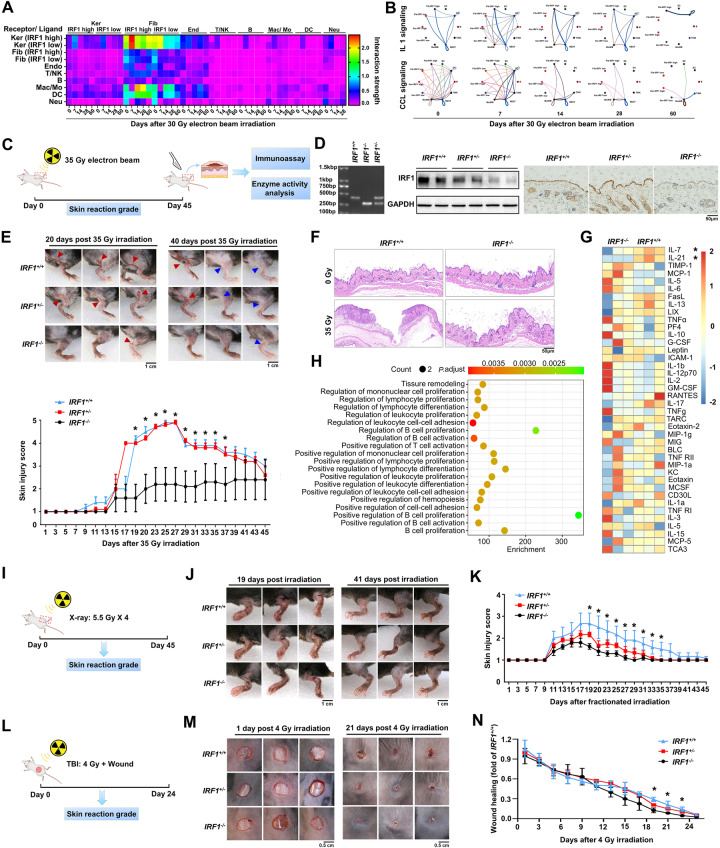
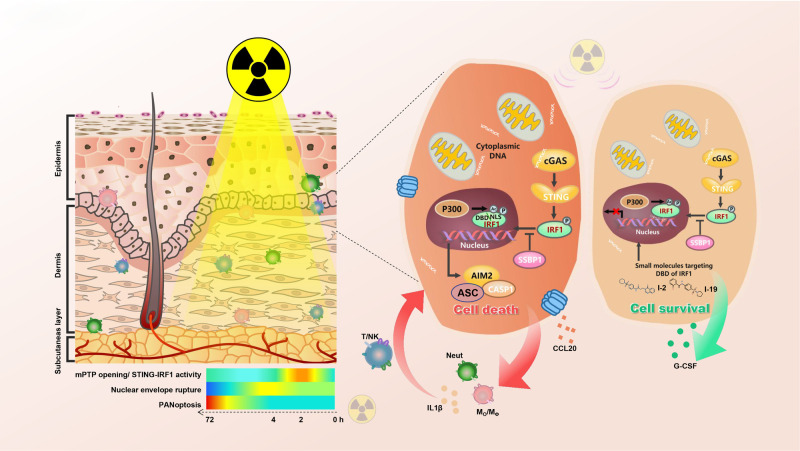
The key role of structural cells in immune modulation has been revealed with the advent of single-cell multiomics, but the underlying mechanism remains poorly understood. Here, we revealed that the transcriptional activation of interferon regulatory factor 1 (IRF1) in response to ionizing radiation, cytotoxic chemicals and SARS-CoV-2 viral infection determines the fate of structural cells and regulates communication between structural and immune cells. Radiation-induced leakage of mtDNA initiates the nuclear translocation of IRF1, enabling it to regulate the transcription of inflammation- and cell death-related genes. Novel posttranslational modification (PTM) sites in the nuclear localization sequence (NLS) of IRF1 were identified. Functional analysis revealed that mutation of the acetylation site and the phosphorylation sites in the NLS blocked the transcriptional activation of IRF1 and reduced cell death in response to ionizing radiation. Mechanistically, reciprocal regulation between the single-stranded DNA sensors SSBP1 and IRF1, which restrains radiation-induced and STING/p300-mediated PTMs of IRF1, was revealed. In addition, genetic deletion or pharmacological inhibition of IRF1 tempered radiation-induced inflammatory cell death, and radiation mitigators also suppressed SARS-CoV-2 NSP-10-mediated activation of IRF1. Thus, we revealed a novel cytoplasm-oriented mechanism of IRF1 activation in structural cells that promotes inflammation and highlighted the potential effectiveness of IRF1 inhibitors against immune disorders.
Keywords: Interferon regulatory factor 1 (IRF1); Ionizing radiation; Nuclear translocation; Posttranslational modification (PTM); Transcription regulation.
© 2024. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
West China Second University Hospital has two patents pending related to the two effective IRF1-responsive inhibitors reported in this work, entitled “A small molecular compound against IRF1-responsive for treating inflammatory skin injury” and “A novel agent against genotoxic-induced tissue injuries”, respectively.
Figures
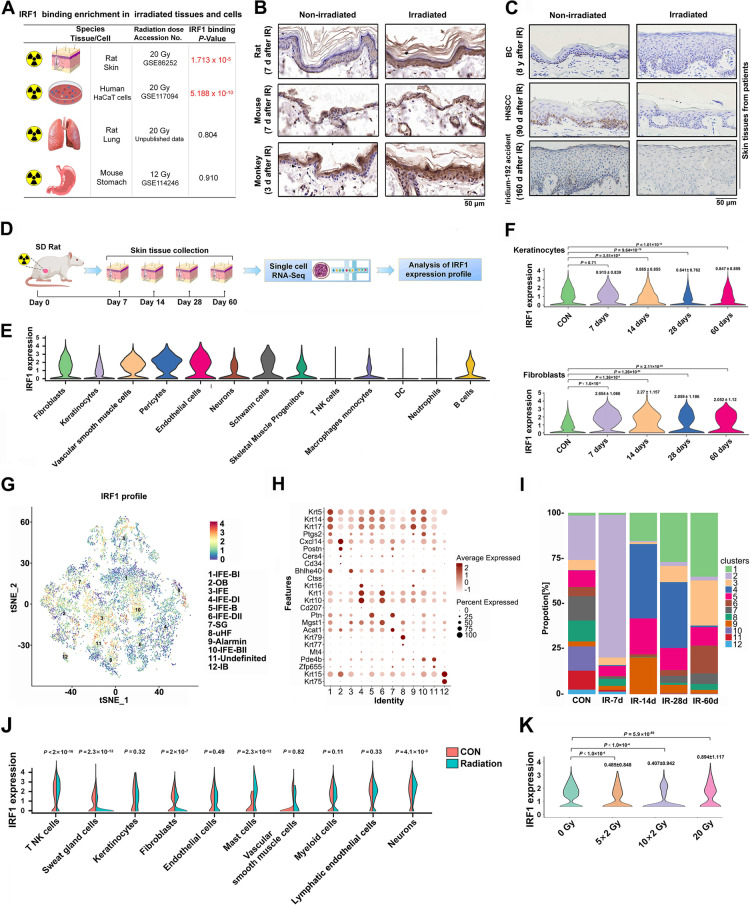
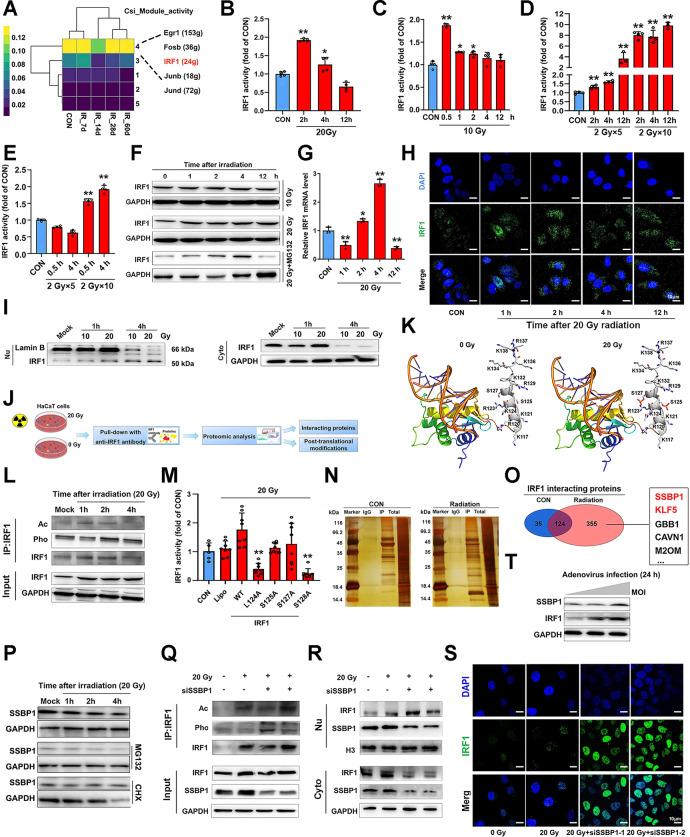
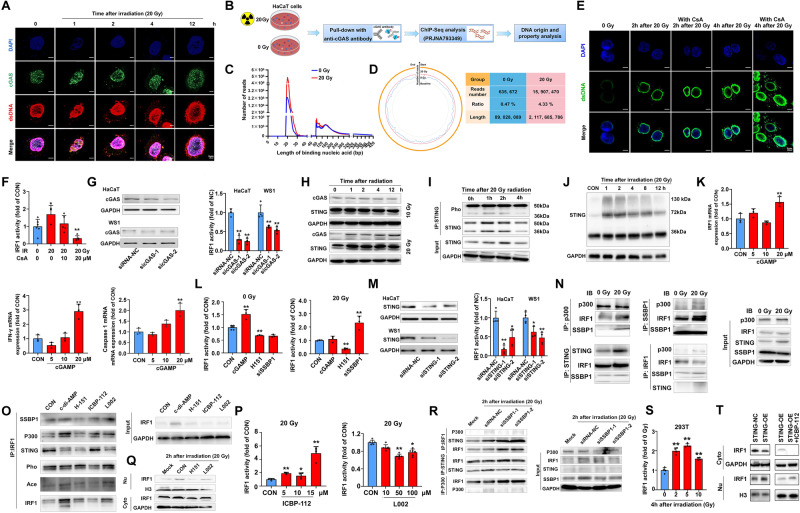
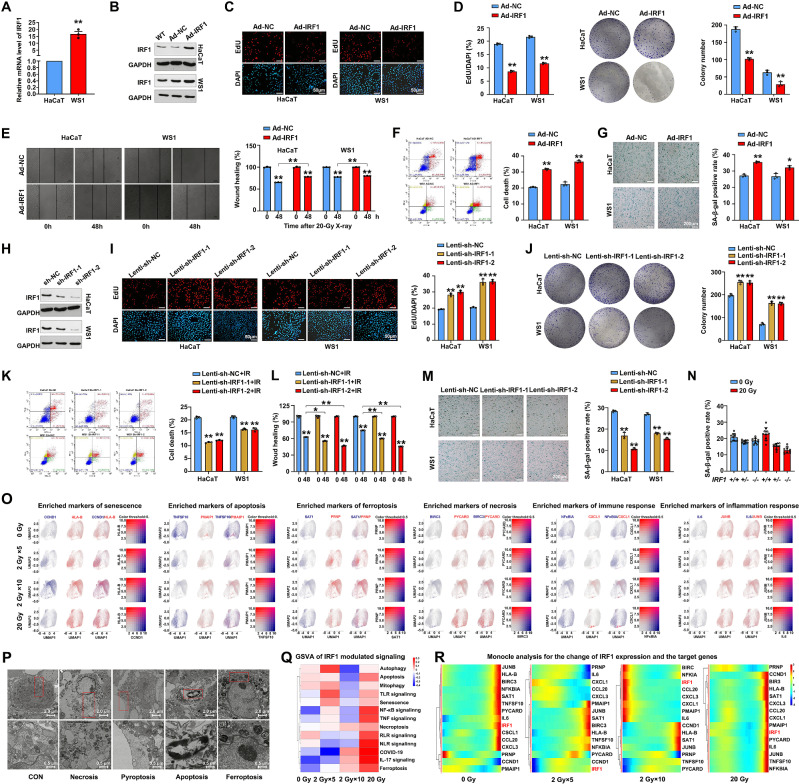


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
