

The FABD domain is critical for the oncogenicity of BCR/ABL in chronic myeloid leukaemia
- PMID: 38849885
- PMCID: PMC11157785
- DOI: 10.1186/s12964-024-01694-8
The FABD domain is critical for the oncogenicity of BCR/ABL in chronic myeloid leukaemia
Abstract
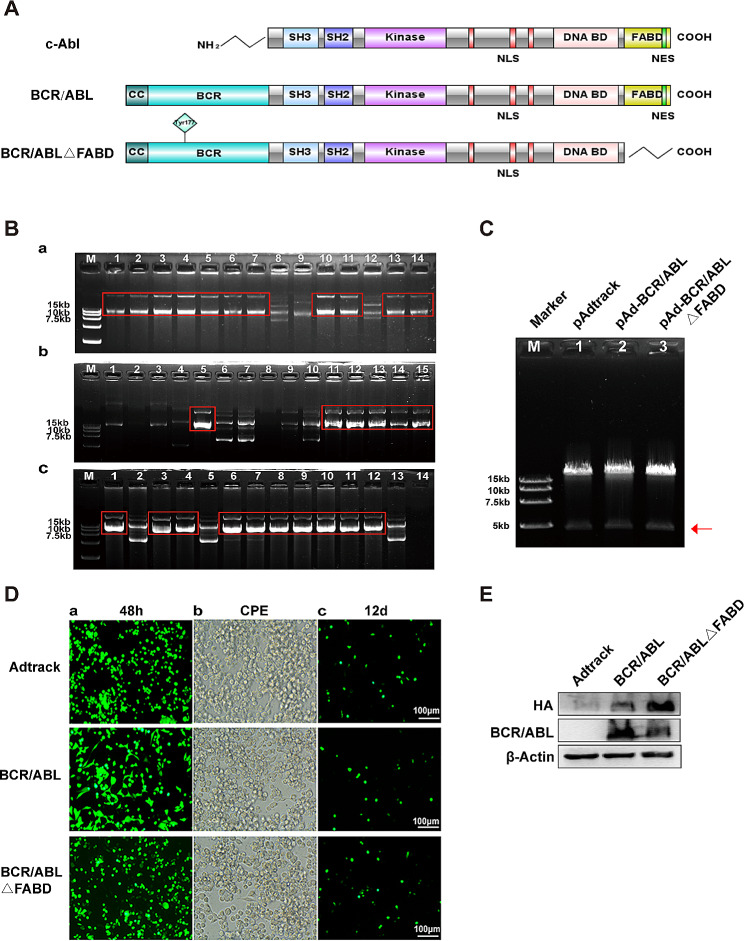
Background: Abnormally expressed BCR/ABL protein serves as the basis for the development of chronic myeloid leukaemia (CML). The F-actin binding domain (FABD), which is a crucial region of the BCR/ABL fusion protein, is also located at the carboxyl end of the c-ABL protein and regulates the kinase activity of c-ABL. However, the precise function of this domain in BCR/ABL remains uncertain.
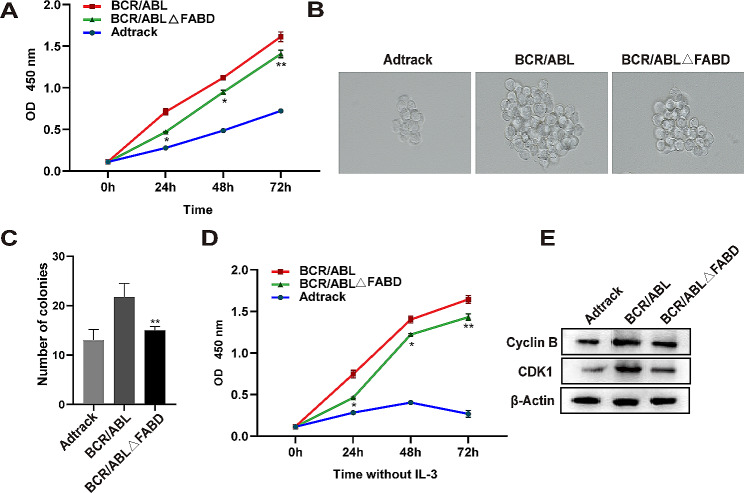
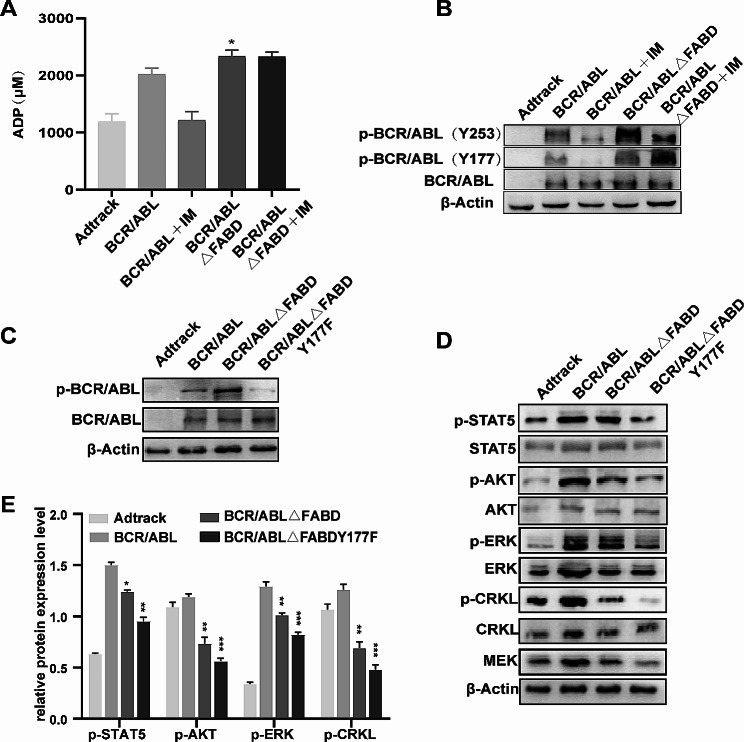
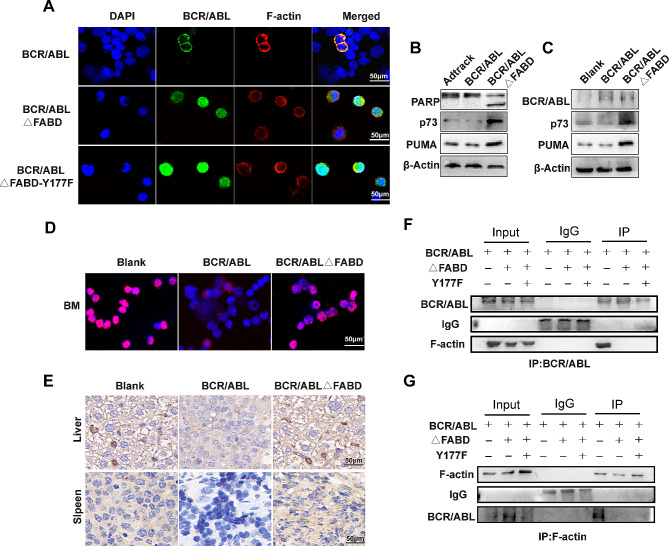
Methods: The FABD-deficient adenovirus vectors Ad-BCR/ABL△FABD, wild-type Ad-BCR/ABL and the control vector Adtrack were constructed, and 32D cells were infected with these adenoviruses separately. The effects of FABD deletion on the proliferation and apoptosis of 32D cells were evaluated by a CCK-8 assay, colony formation assay, flow cytometry and DAPI staining. The levels of phosphorylated BCR/ABL, p73, and their downstream signalling molecules were detected by western blot. The intracellular localization and interaction of BCR/ABL with the cytoskeleton-related protein F-actin were identified by immunofluorescence and co-IP. The effect of FABD deletion on BCR/ABL carcinogenesis in vivo was explored in CML-like mouse models. The degree of leukaemic cell infiltration was observed by Wright‒Giemsa staining and haematoxylin and eosin (HE) staining.
Results: We report that the loss of FABD weakened the proliferation-promoting ability of BCR/ABL, accompanied by the downregulation of BCR/ABL downstream signals. Moreover, the deletion of FABD resulted in a change in the localization of BCR/ABL from the cytoplasm to the nucleus, accompanied by an increase in cell apoptosis due to the upregulation of p73 and its downstream proapoptotic factors. Furthermore, we discovered that the absence of FABD alleviated leukaemic cell infiltration induced by BCR/ABL in mice.
Conclusions: These findings reveal that the deletion of FABD diminished the carcinogenic potential of BCR/ABL both in vitro and in vivo. This study provides further insight into the function of the FABD domain in BCR/ABL.
Keywords: BCR/ABL; Chronic myeloid leukaemia; F-actin binding domain; Oncogenicity; p73.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
