

iSuRe-HadCre is an essential tool for effective conditional genetics
- PMID: 38850155
- PMCID: PMC11260470
- DOI: 10.1093/nar/gkae472
iSuRe-HadCre is an essential tool for effective conditional genetics
Abstract
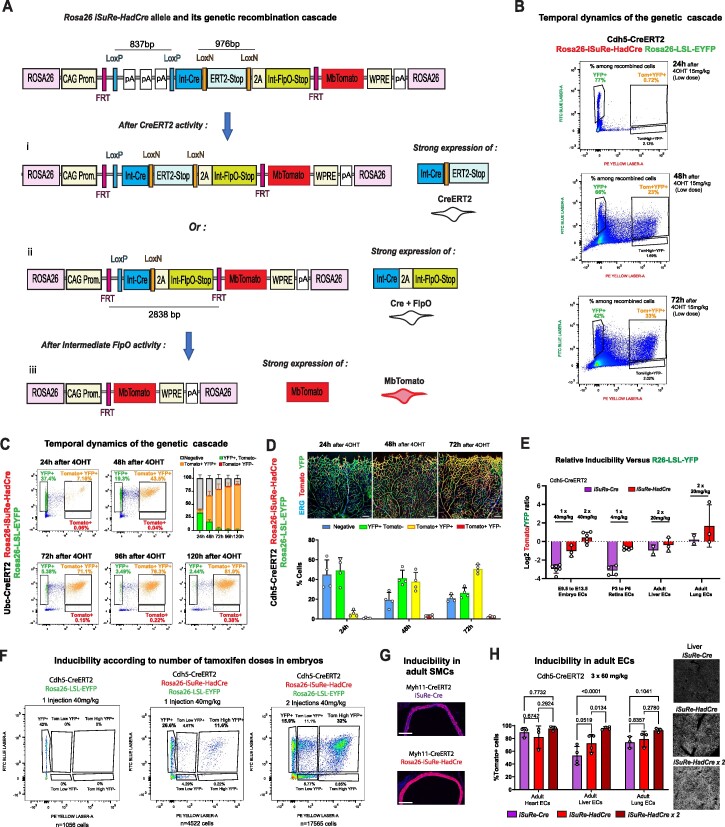
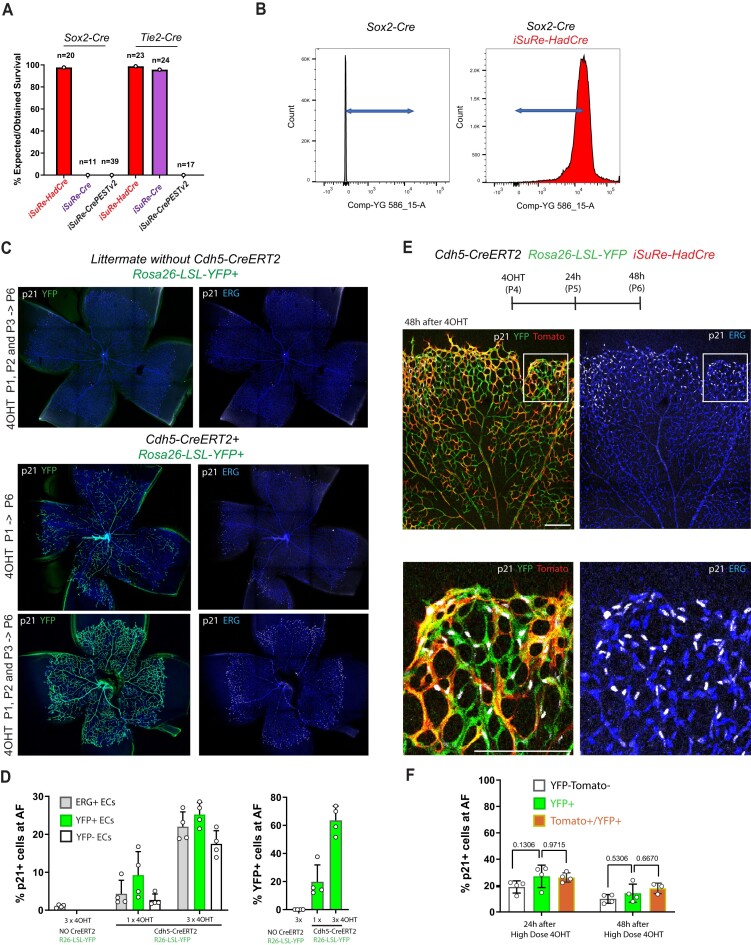
Methods for modifying gene function at high spatiotemporal resolution in mice have revolutionized biomedical research, with Cre-loxP being the most widely used technology. However, the Cre-loxP technology has several drawbacks, including weak activity, leakiness, toxicity, and low reliability of existing Cre-reporters. This is mainly because different genes flanked by loxP sites (floxed) vary widely in their sensitivity to Cre-mediated recombination. Here, we report the generation, validation, and utility of iSuRe-HadCre, a new dual Cre-reporter and deleter mouse line that avoids these drawbacks. iSuRe-HadCre achieves this through a novel inducible dual-recombinase genetic cascade that ensures that cells expressing a fluorescent reporter had only transient Cre activity, that is nonetheless sufficient to effectively delete floxed genes. iSuRe-HadCre worked reliably in all cell types and for the 13 floxed genes tested. This new tool will enable the precise, efficient, and trustworthy analysis of gene function in entire mouse tissues or in single cells.
© The Author(s) 2024. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures








References
-
- Luo L., Ambrozkiewicz M.C., Benseler F., Chen C., Dumontier E., Falkner S., Furlanis E., Gomez A.M., Hoshina N., Huang W.H. et al. Optimizing Nervous System-Specific Gene Targeting with Cre Driver Lines: Prevalence of Germline Recombination and Influencing Factors. Neuron. 2020; 106:37–65. - PMC - PubMed
-
- Schmidt-Supprian M., Rajewsky K. Vagaries of conditional gene targeting. Nat. Immunol. 2007; 8:665–668. - PubMed
MeSH terms
Substances
Grants and funding
- PI R.B
- MCIN - PID2020-120252RB-I00/Ministerio de Ciencia e Innovación
- HR19-00120/Fundacion La Caixa
- 2018.0218/Knut and Alice Wallenberg Foundation
- Göran Gustafsson foundation
- CX-SO-16-1/Fundación La Caixa
- Swedish Society for Medical Research
- European Union's Horizon 2020
- 814316/Marie Skłodowska-Curie
- ReDIB ICTS infrastructure TRIMA@CNIC
- Ministerio de Ciencia e Innovación
- Instituto de Salud Carlos III
- Pro CNIC Foundation
- CEX2020-001041-S/Severo Ochoa Center of Excellence
- ERC-2020-COG- 101001814/ERC_/European Research Council/International
- EMERGE-773047/ERC_/European Research Council/International
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
