

Atlastin-1 regulates endosomal tubulation and lysosomal proteolysis in human cortical neurons
- PMID: 38851544
- PMCID: PMC11300884
- DOI: 10.1016/j.nbd.2024.106556
Atlastin-1 regulates endosomal tubulation and lysosomal proteolysis in human cortical neurons
Abstract
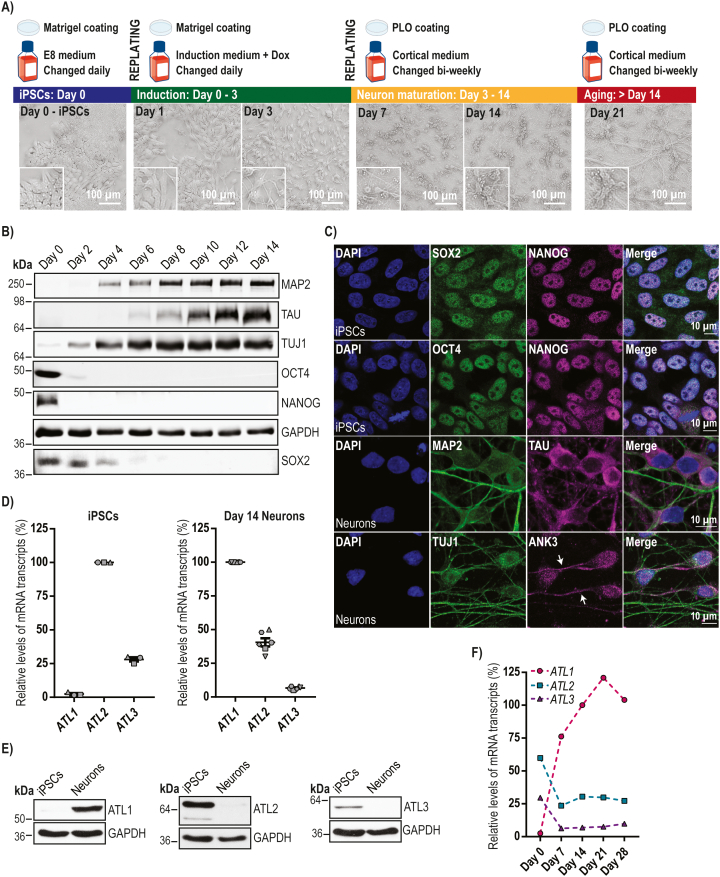
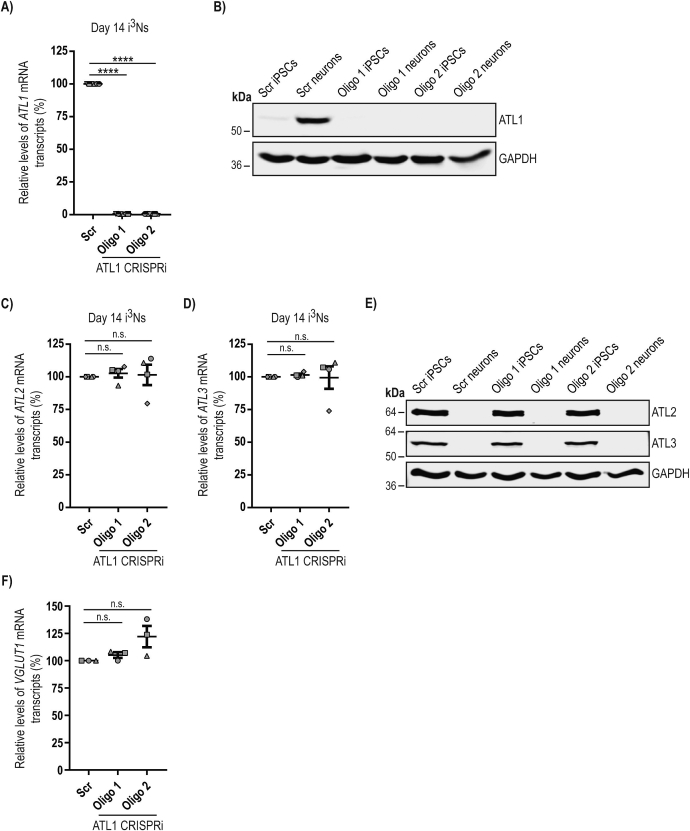
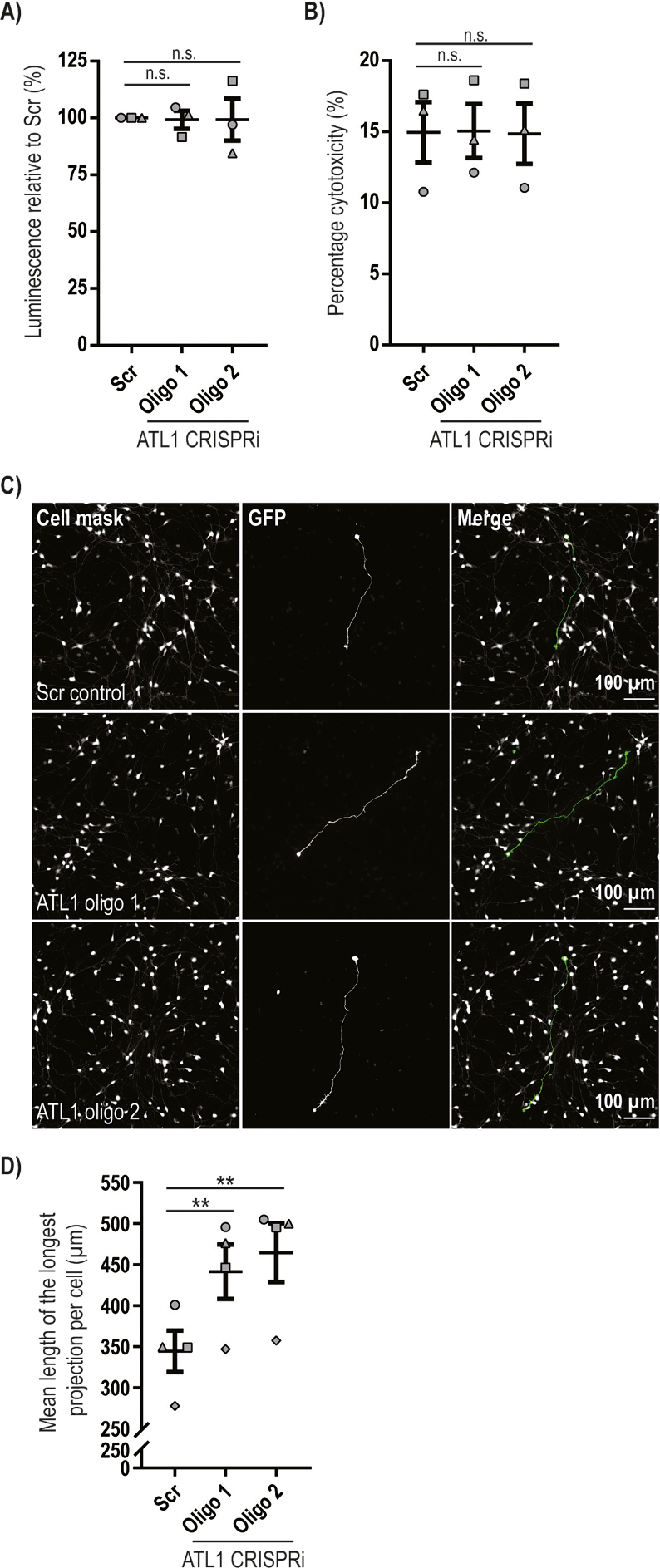
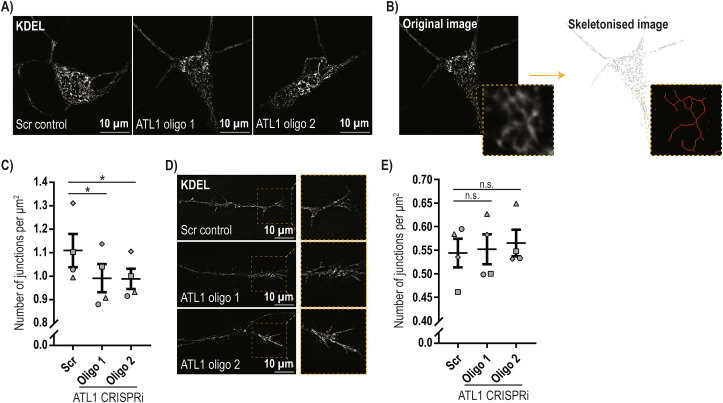
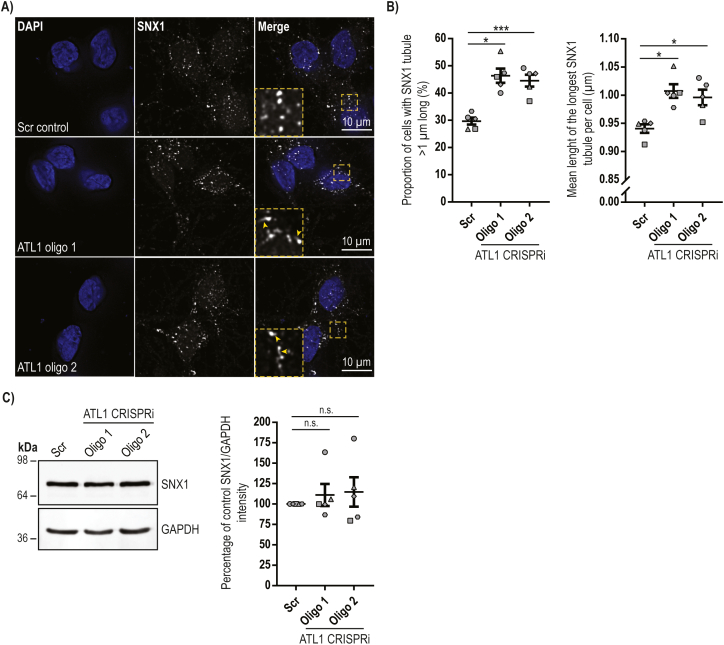
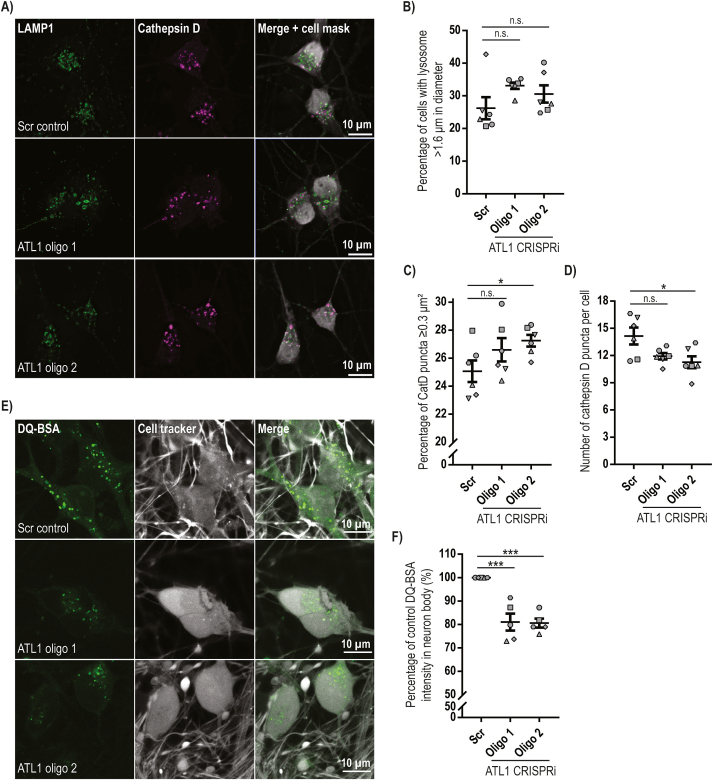
Mutation of the ATL1 gene is one of the most common causes of hereditary spastic paraplegia (HSP), a group of genetic neurodegenerative conditions characterised by distal axonal degeneration of the corticospinal tract axons. Atlastin-1, the protein encoded by ATL1, is one of three mammalian atlastins, which are homologous dynamin-like GTPases that control endoplasmic reticulum (ER) morphology by fusing tubules to form the three-way junctions that characterise ER networks. However, it is not clear whether atlastin-1 is required for correct ER morphology in human neurons and if so what the functional consequences of lack of atlastin-1 are. Using CRISPR-inhibition we generated human cortical neurons lacking atlastin-1. We demonstrate that ER morphology was altered in these neurons, with a reduced number of three-way junctions. Neurons lacking atlastin-1 had longer endosomal tubules, suggestive of defective tubule fission. This was accompanied by reduced lysosomal proteolytic capacity. As well as demonstrating that atlastin-1 is required for correct ER morphology in human neurons, our results indicate that lack of a classical ER-shaping protein such as atlastin-1 may cause altered endosomal tubulation and lysosomal proteolytic dysfunction. Furthermore, they strengthen the idea that defective lysosome function contributes to the pathogenesis of a broad group of HSPs, including those where the primary localisation of the protein involved is not at the endolysosomal system.
Keywords: Atlastin; Endoplasmic reticulum morphology; Endosomal traffic; Endosomal tubulation; Hereditary spastic paraplegia; Lysosomal proteolysis; Lysosome.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Evan Reid reports a relationship with SwanBio Therapeutics, Inc. that includes: consulting or advisory. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Blackstone C. Hereditary spastic paraplegia. Handb. Clin. Neurol. 2018;148:633–652. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
