

Crosslinking of base-modified RNAs by synthetic DYW-KP base editors implicates an enzymatic lysine as the nitrogen donor for U-to-C RNA editing
- PMID: 38852885
- PMCID: PMC11332814
- DOI: 10.1016/j.jbc.2024.107454
Crosslinking of base-modified RNAs by synthetic DYW-KP base editors implicates an enzymatic lysine as the nitrogen donor for U-to-C RNA editing
Abstract
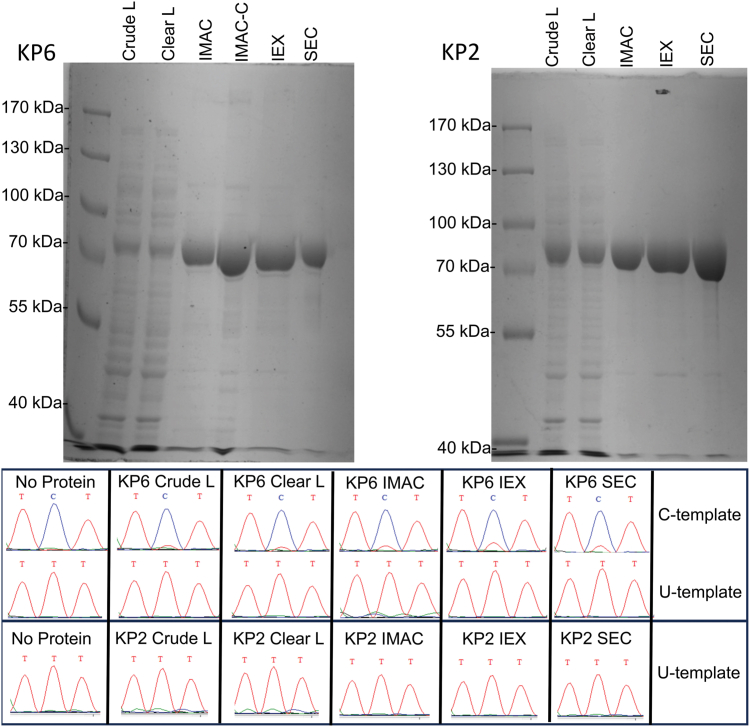
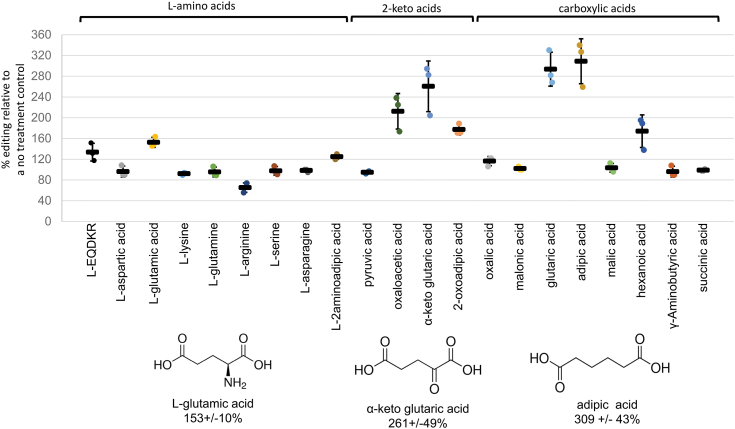
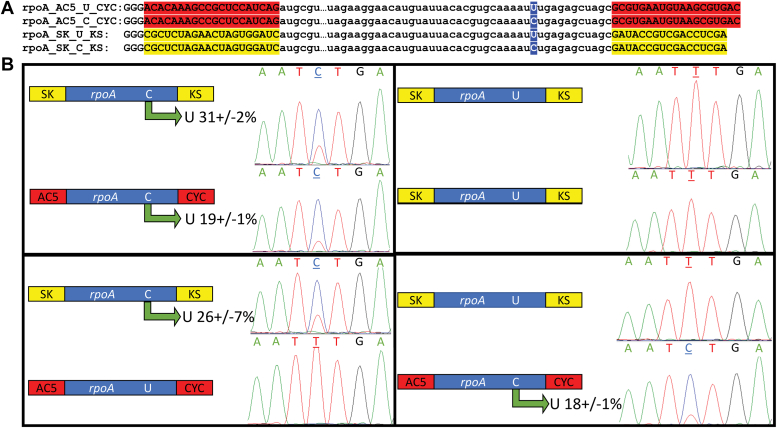
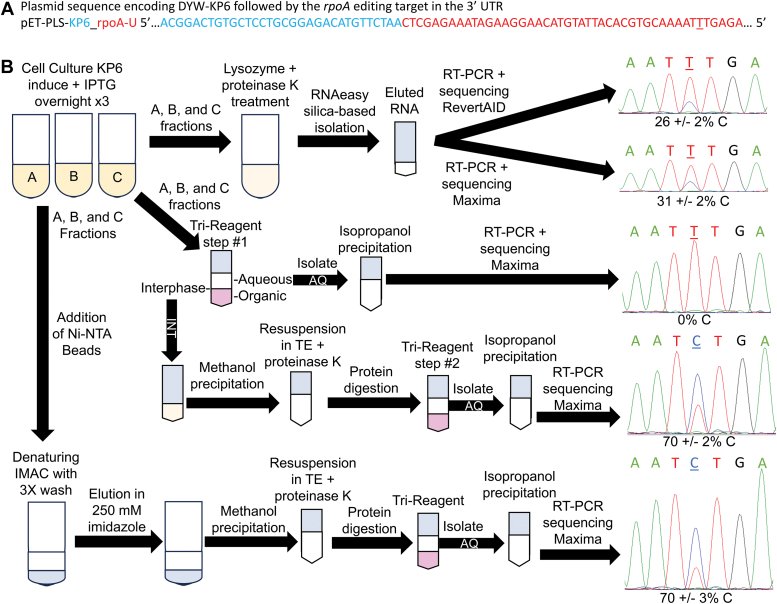
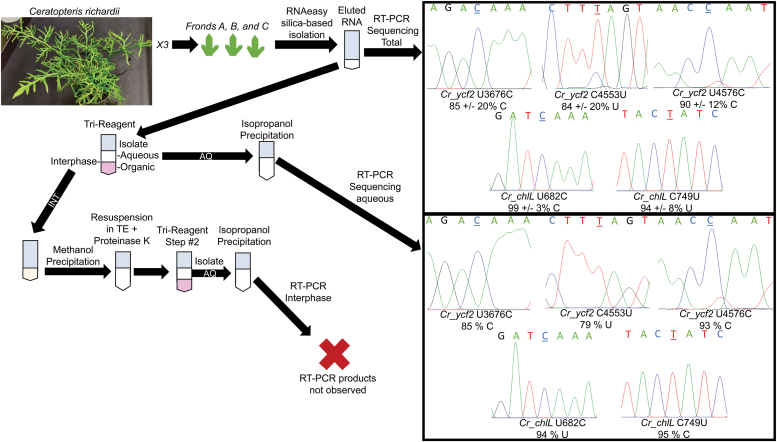
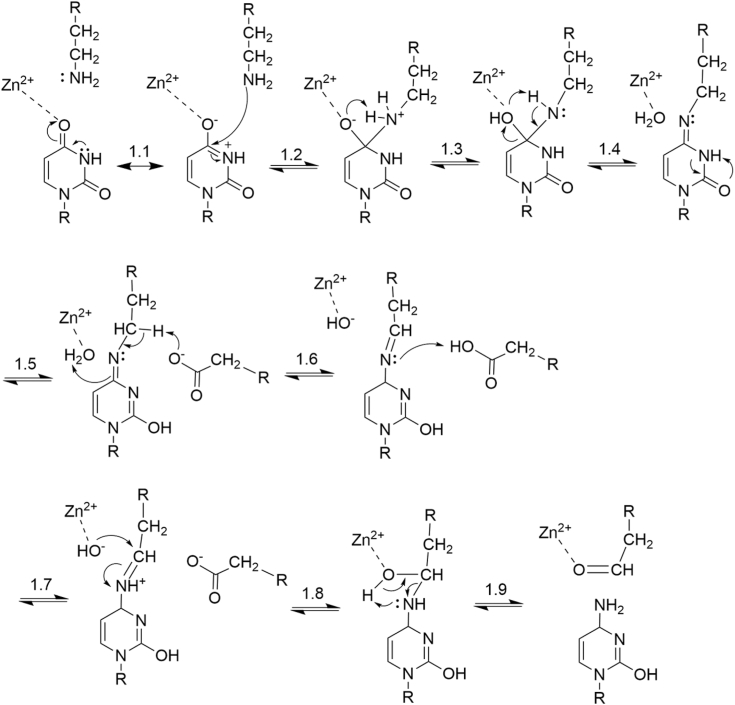
Sequence-specific cytidine to uridine (C-to-U) and adenosine to inosine editing tools can alter RNA and DNA sequences and utilize a hydrolytic deamination mechanism requiring an active site zinc ion and a glutamate residue. In plant organelles, DYW-PG domain containing enzymes catalyze C-to-U edits through the canonical deamination mechanism. Proteins developed from consensus sequences of the related DYW-KP domain family catalyze what initially appeared to be uridine to cytidine (U-to-C) edits leading to this investigation into the U-to-C editing mechanism. The synthetic DYW-KP enzyme KP6 was found sufficient for C-to-U editing activity stimulated by the addition of carboxylic acids in vitro. Despite addition of putative amine/amide donors, U-to-C editing by KP6 could not be observed in vitro. C-to-U editing was found not to be concomitant with U-to-C editing, discounting a pyrimidine transaminase mechanism. RNAs containing base modifications were highly enriched in interphase fractions consistent with covalent crosslinks to KP6, KP2, and KP3 proteins. Mass spectrometry of purified KP2 and KP6 proteins revealed secondary peaks with mass shifts of 319 Da. A U-to-C crosslinking mechanism was projected to explain the link between crosslinking, RNA base changes, and the ∼319 Da mass. In this model, an enzymatic lysine attacks C4 of uridine to form a Schiff base RNA-protein conjugate. Sequenced RT-PCR products from the fern Ceratopteris richardii indicate U-to-C base edits do not preserve proteinaceous crosslinks in planta. Hydrolysis of a protonated Schiff base conjugate releasing cytidine is hypothesized to explain the completed pathway in plants.
Keywords: C-to-U RNA editing; Ceratopteris richardii; RNA crosslinking; U-to-C RNA editing; metalloenzymes.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interests The authors declare no conflicts of interest with the contents of this article.
Figures
Similar articles
-
A Conserved Lysine in an Ion-Pair with a Catalytic Glutamate Is Critical for U-to-C RNA Editing but Restricts C-to-U RNA Editing.Biochemistry. 2025 Jan 7;64(1):15-19. doi: 10.1021/acs.biochem.4c00625. Epub 2024 Dec 9. Biochemistry. 2025. PMID: 39653594 Free PMC article.
-
Fine-tuning of the PPR protein directs the RNA editing activity toward C-to-U or U-to-C conversion.Sci Rep. 2025 Feb 21;15(1):6288. doi: 10.1038/s41598-025-90722-9. Sci Rep. 2025. PMID: 39984571 Free PMC article.
-
U-to-C RNA editing by synthetic PPR-DYW proteins in bacteria and human culture cells.Commun Biol. 2022 Sep 15;5(1):968. doi: 10.1038/s42003-022-03927-3. Commun Biol. 2022. PMID: 36109586 Free PMC article.
-
C-to-U editing and site-directed RNA editing for the correction of genetic mutations.Biosci Trends. 2017 Jul 24;11(3):243-253. doi: 10.5582/bst.2017.01049. Epub 2017 May 8. Biosci Trends. 2017. PMID: 28484188 Review.
-
RNA editing in plant mitochondria—connecting RNA target sequences and acting proteins.Mitochondrion. 2014 Nov;19 Pt B:191-7. doi: 10.1016/j.mito.2014.04.005. Epub 2014 Apr 13. Mitochondrion. 2014. PMID: 24732437 Review.
Cited by
-
A Conserved Lysine in an Ion-Pair with a Catalytic Glutamate Is Critical for U-to-C RNA Editing but Restricts C-to-U RNA Editing.Biochemistry. 2025 Jan 7;64(1):15-19. doi: 10.1021/acs.biochem.4c00625. Epub 2024 Dec 9. Biochemistry. 2025. PMID: 39653594 Free PMC article.
-
Fine-tuning of the PPR protein directs the RNA editing activity toward C-to-U or U-to-C conversion.Sci Rep. 2025 Feb 21;15(1):6288. doi: 10.1038/s41598-025-90722-9. Sci Rep. 2025. PMID: 39984571 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
