

This is a preprint.
Neuronal segmentation in cephalopod arms
- PMID: 38853825
- PMCID: PMC11160704
- DOI: 10.1101/2024.05.29.596333
Neuronal segmentation in cephalopod arms
Update in
-
Neuronal segmentation in cephalopod arms.Nat Commun. 2025 Jan 15;16(1):443. doi: 10.1038/s41467-024-55475-5. Nat Commun. 2025. PMID: 39814765 Free PMC article.
Abstract
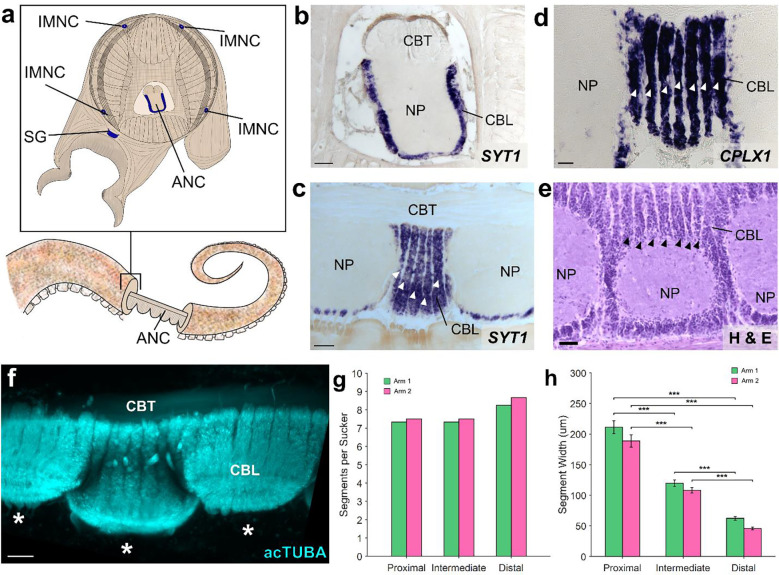
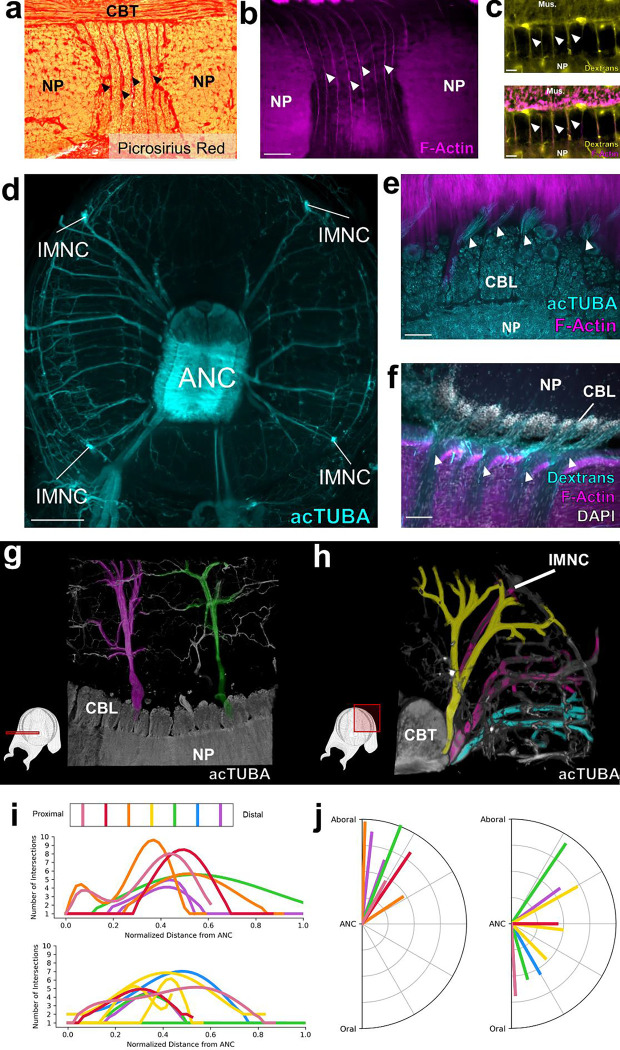
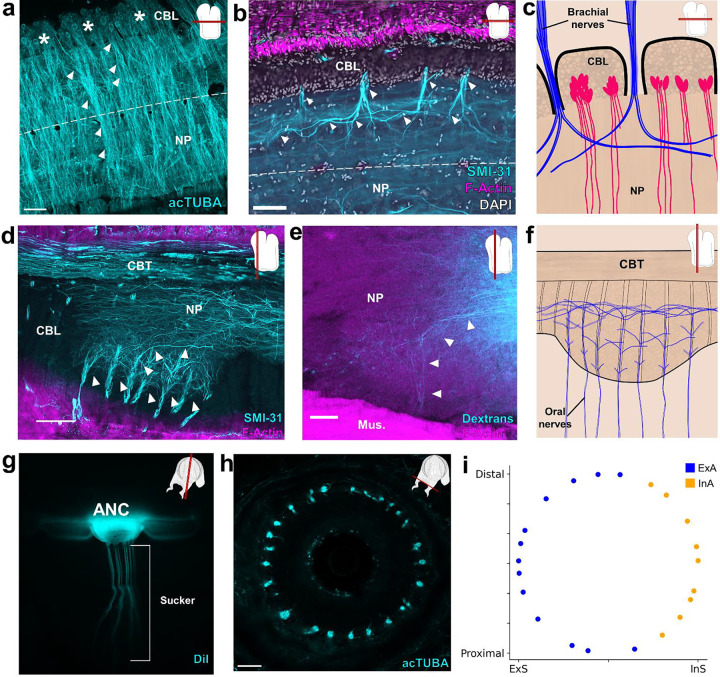
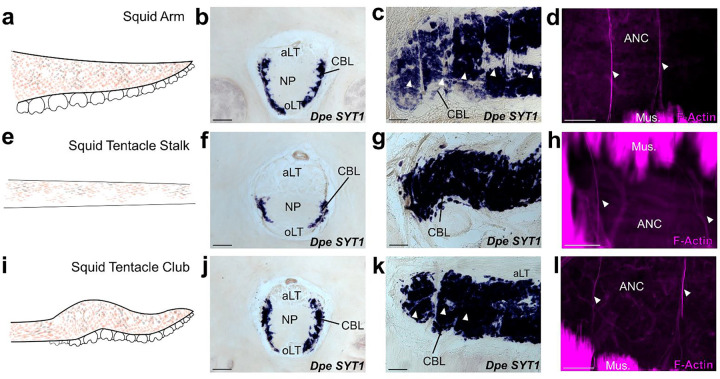
The prehensile arms of the cephalopod are among these animals most remarkable features, but the neural circuitry governing arm and sucker movements remains largely unknown. We studied the neuronal organization of the adult axial nerve cord (ANC) of Octopus bimaculoides with molecular and cellular methods. The ANCs, which lie in the center of every arm, are the largest neuronal structures in the octopus, containing four times as many neurons as found in the central brain. In transverse cross section, the cell body layer (CBL) of the ANC wraps around its neuropil (NP) with little apparent segregation of sensory and motor neurons or nerve exits. Strikingly, when studied in longitudinal sections, the ANC is segmented. ANC neuronal cell bodies form columns separated by septa, with 15 segments overlying each pair of suckers. The segments underlie a modular organization to the ANC neuropil: neuronal cell bodies within each segment send the bulk of their processes directly into the adjoining neuropil, with some reaching the contralateral side. In addition, some nerve processes branch upon entering the NP, forming short-range projections to neighboring segments and mid-range projections to the ANC segments of adjoining suckers. The septa between the segments are employed as ANC nerve exits and as channels for ANC vasculature. Cellular analysis establishes that adjoining septa issue nerves with distinct fiber trajectories, which across two segments (or three septa) fully innervate the arm musculature. Sucker nerves also use the septa, setting up a nerve fiber "suckerotopy" in the sucker-side of the ANC. Comparative anatomy suggests a strong link between segmentation and flexible sucker-laden arms. In the squid Doryteuthis pealeii, the arms and the sucker-rich club of the tentacles have segments, but the sucker-poor stalk of the tentacles does not. The neural modules described here provide a new template for understanding the motor control of octopus soft tissues. In addition, this finding represents the first demonstration of nervous system segmentation in a mollusc.
Figures
Similar articles
-
Neuronal segmentation in cephalopod arms.Res Sq [Preprint]. 2024 Jul 4:rs.3.rs-4548192. doi: 10.21203/rs.3.rs-4548192/v1. Res Sq. 2024. Update in: Nat Commun. 2025 Jan 15;16(1):443. doi: 10.1038/s41467-024-55475-5. PMID: 39011093 Free PMC article. Updated. Preprint.
-
Neuronal segmentation in cephalopod arms.Nat Commun. 2025 Jan 15;16(1):443. doi: 10.1038/s41467-024-55475-5. Nat Commun. 2025. PMID: 39814765 Free PMC article.
-
Molecular and morphological circuitry of the octopus sucker ganglion.bioRxiv [Preprint]. 2025 Feb 11:2025.02.10.637560. doi: 10.1101/2025.02.10.637560. bioRxiv. 2025. Update in: J Comp Neurol. 2025 May;533(5):e70055. doi: 10.1002/cne.70055. PMID: 39990388 Free PMC article. Updated. Preprint.
-
Inspiration, simulation and design for smart robot manipulators from the sucker actuation mechanism of cephalopods.Bioinspir Biomim. 2007 Dec;2(4):S170-81. doi: 10.1088/1748-3182/2/4/S06. Epub 2007 Oct 16. Bioinspir Biomim. 2007. PMID: 18037726 Review.
-
Toward an Understanding of Octopus Arm Motor Control.Integr Comp Biol. 2023 Dec 29;63(6):1277-1284. doi: 10.1093/icb/icad069. Integr Comp Biol. 2023. PMID: 37327080 Free PMC article. Review.
References
-
- Hochner B., Zullo L., Shomrat T., Levy G. & Nesher N. Embodied mechanisms of motor control in the octopus. Curr. Biol. 33, R1119–R1125 (2023). - PubMed
-
- Kier W. M. & Stella M. P. The arrangement and function of octopus arm musculature and connective tissue. J. Morphol. 268, 831–843 (2007). - PubMed
-
- Grasso F. W. Octopus sucker-arm coordination in grasping and manipulation*. Am. Malacol. Bull. 24, 13–23 (2008).
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous