

This is a preprint.
The context-dependent epigenetic and organogenesis programs determine 3D vs. 2D cellular fitness of MYC-driven murine liver cancer cells
- PMID: 38853928
- PMCID: PMC11160912
- DOI: 10.21203/rs.3.rs-4390765/v4
The context-dependent epigenetic and organogenesis programs determine 3D vs. 2D cellular fitness of MYC-driven murine liver cancer cells
Update in
-
The context-dependent epigenetic and organogenesis programs determine 3D vs. 2D cellular fitness of MYC-driven murine liver cancer cells.Elife. 2025 May 6;14:RP101299. doi: 10.7554/eLife.101299. Elife. 2025. PMID: 40326560 Free PMC article.
Abstract
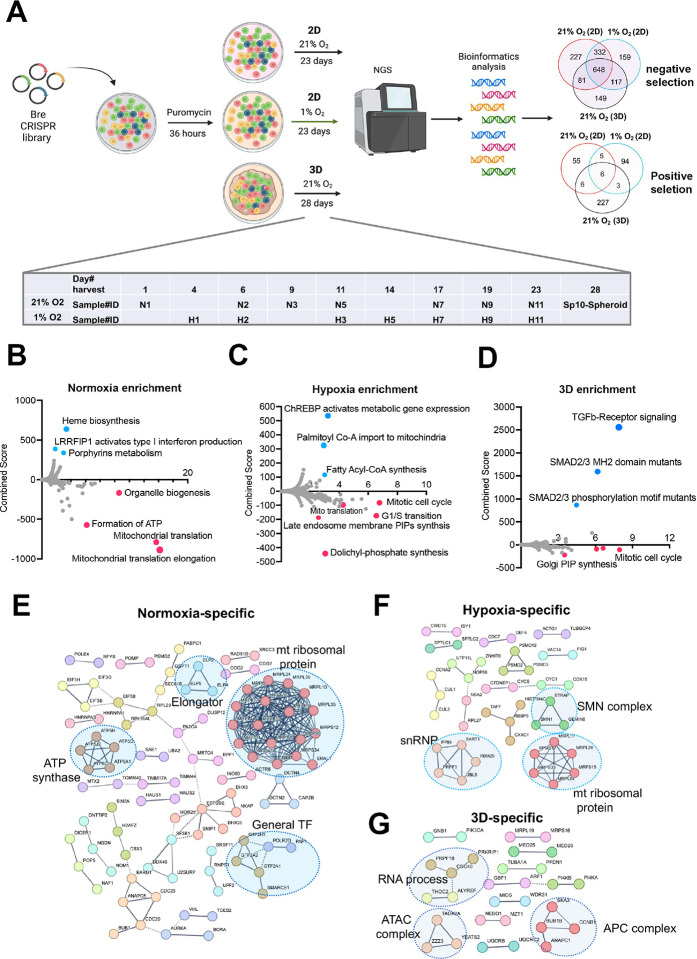
3D cellular-specific epigenetic and transcriptomic reprogramming is critical to organogenesis and tumorigenesis. Here we dissect the distinct cell fitness in 2D (normoxia vs. chronic hypoxia) vs 3D (normoxia) culture conditions for a MYC-driven murine liver cancer model. We identify over 600 shared essential genes and additional context-specific fitness genes and pathways. Knockout of the VHL-HIF1 pathway results in incompatible fitness defects under normoxia vs. 1% oxygen or 3D culture conditions. Moreover, deletion of each of the mitochondrial respiratory electron transport chain complex has distinct fitness outcomes. Notably, multicellular organogenesis signaling pathways including TGFb-SMAD, which is upregulated in 3D culture, specifically constrict the uncontrolled cell proliferation in 3D while inactivation of epigenetic modifiers (Bcor, Kmt2d, METTL3 and METTL14) has opposite outcomes in 2D vs. 3D. We further identify a 3D-dependent synthetic lethality with partial loss of Prmt5 due to a reduction of Mtap expression resulting from 3D-specific epigenetic reprogramming. Our study highlights unique epigenetic, metabolic and organogenesis signaling dependencies under different cellular settings.
Figures






Similar articles
-
The context-dependent epigenetic and organogenesis programs determine 3D vs. 2D cellular fitness of MYC-driven murine liver cancer cells.Elife. 2025 May 6;14:RP101299. doi: 10.7554/eLife.101299. Elife. 2025. PMID: 40326560 Free PMC article.
-
A Hybrid 2D/3D Approach for Neural Differentiation Into Telencephalic Organoids and Efficient Modulation of FGF8 Signaling.Bio Protoc. 2025 Jun 20;15(12):e5354. doi: 10.21769/BioProtoc.5354. eCollection 2025 Jun 20. Bio Protoc. 2025. PMID: 40620811 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
[Volume and health outcomes: evidence from systematic reviews and from evaluation of Italian hospital data].Epidemiol Prev. 2013 Mar-Jun;37(2-3 Suppl 2):1-100. Epidemiol Prev. 2013. PMID: 23851286 Italian.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
References
-
- McDonald ER 3rd, de Weck A, Schlabach MR, Billy E, Mavrakis KJ, Hoffman GR, Belur D, Castelletti D, Frias E, Gampa K et al. (2017) Project DRIVE: A Compendium of Cancer Dependencies and Synthetic Lethal Relationships Uncovered by Large-Scale. Deep RNAi Screen Cell 170:577–592e510. 10.1016/j.cell.2017.07.005 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
