

This is a preprint.
Single-nucleus multi-omic profiling of polyploid heart nuclei identifies fusion-derived cardiomyocytes in the human heart
- PMID: 38853931
- PMCID: PMC11160865
- DOI: 10.21203/rs.3.rs-4414468/v1
Single-nucleus multi-omic profiling of polyploid heart nuclei identifies fusion-derived cardiomyocytes in the human heart
Abstract
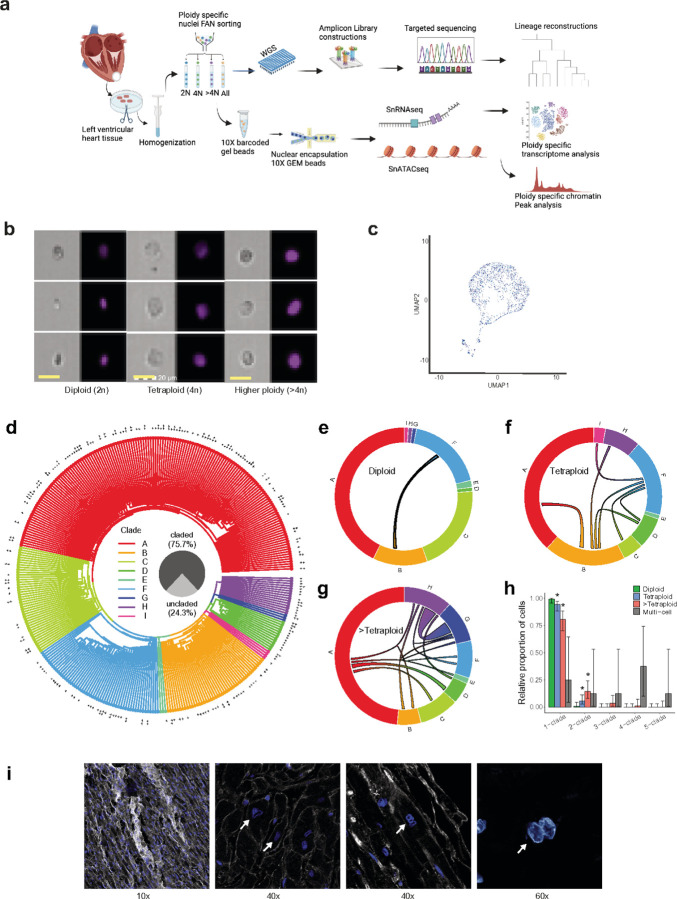
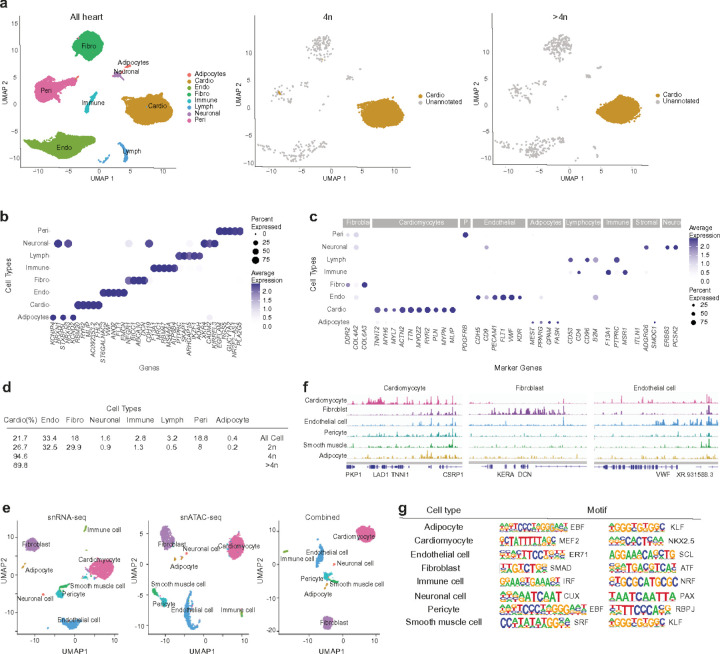
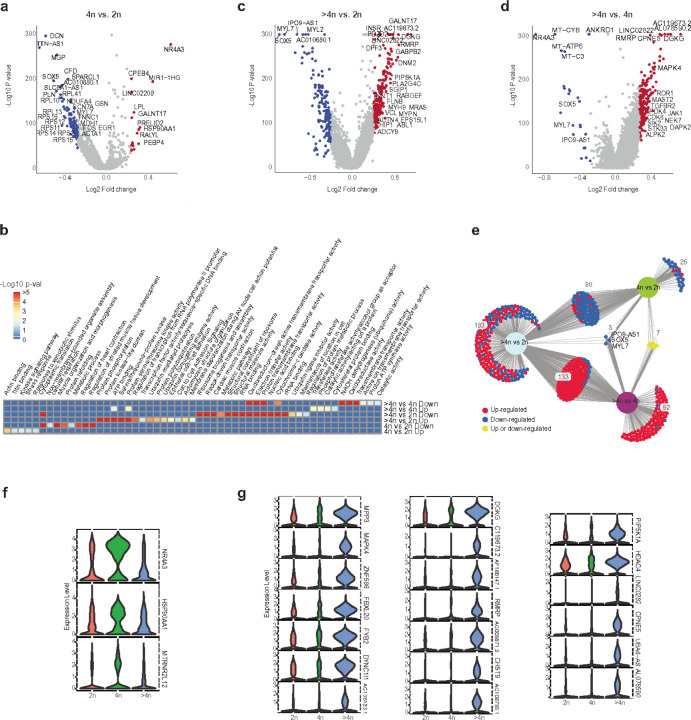
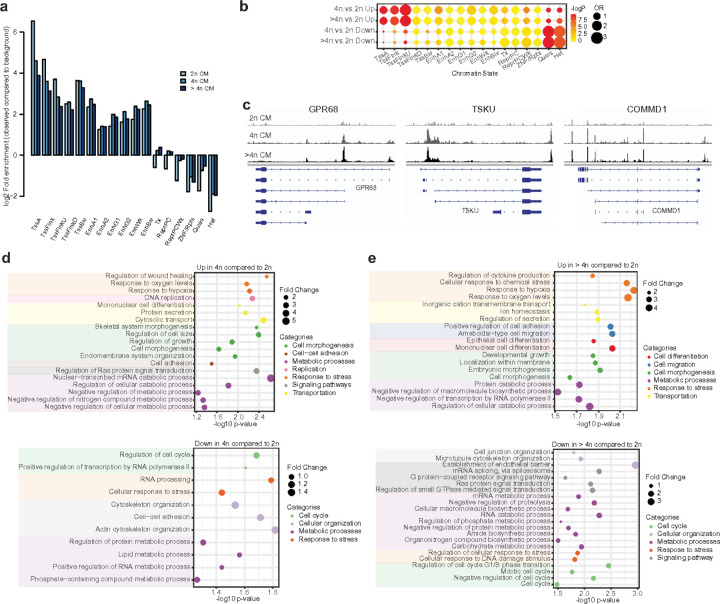
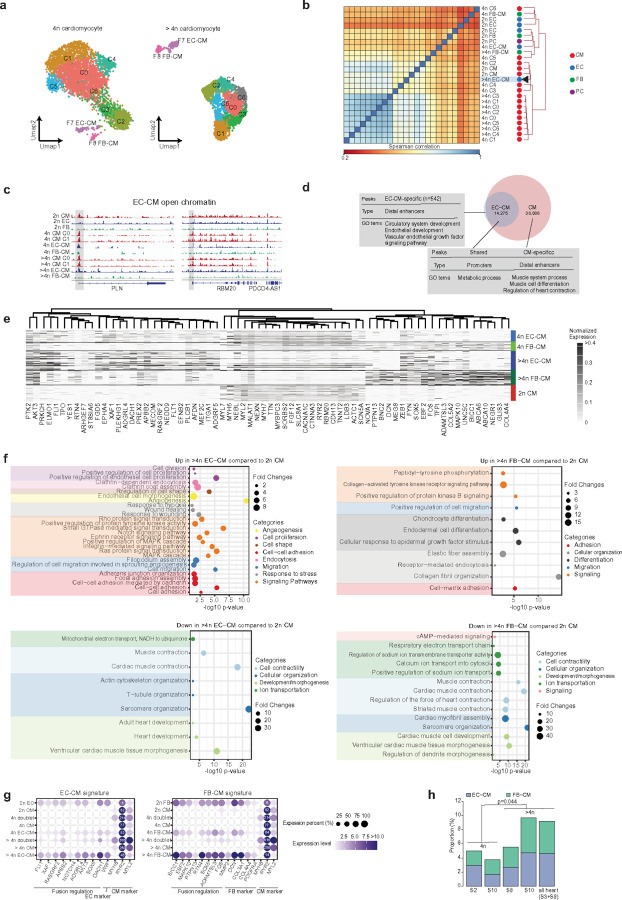
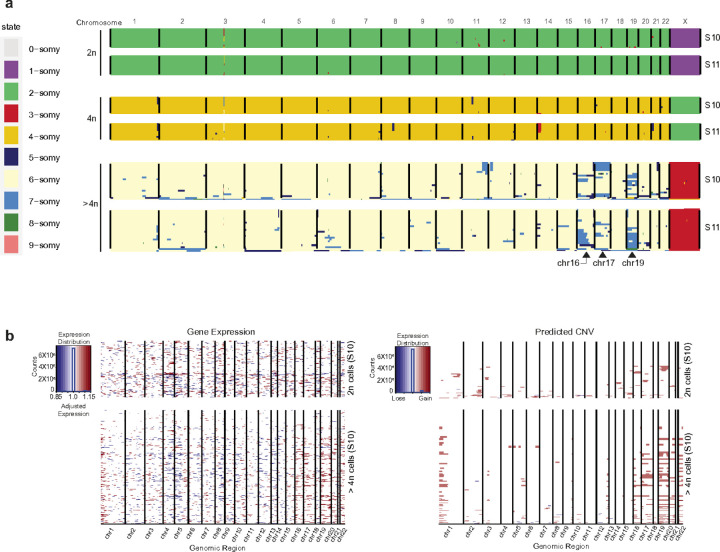
Understanding the mechanisms of polyploidization in cardiomyocytes is crucial for advancing strategies to stimulate myocardial regeneration. Although endoreplication has long been considered the primary source of polyploid human cardiomyocytes, recent animal work suggests the potential for cardiomyocyte fusion. Moreover, the effects of polyploidization on the genomic-transcriptomic repertoire of human cardiomyocytes have not been studied previously. We applied single-nuclei whole genome sequencing, single nuclei RNA sequencing, and multiome ATAC + gene expression (from the same nuclei) techniques to nuclei isolated from 11 healthy hearts. Utilizing post-zygotic non-inherited somatic mutations occurring during development as "endogenous barcodes," to reconstruct lineage relationships of polyploid cardiomyocytes. Of 482 cardiomyocytes from multiple healthy donor hearts 75.7% can be sorted into several developmental clades marked by one or more somatic single-nucleotide variants (SNVs). At least ~10% of tetraploid cardiomyocytes contain cells from distinct clades, indicating fusion of lineally distinct cells, whereas 60% of higher-ploidy cardiomyocytes contain fused cells from distinct clades. Combined snRNA-seq and snATAC-seq revealed transcriptome and chromatin landscapes of polyploid cardiomyocytes distinct from diploid cardiomyocytes, and show some higher-ploidy cardiomyocytes with transcriptional signatures suggesting fusion between cardiomyocytes and endothelial and fibroblast cells. These observations provide the first evidence for cell and nuclear fusion of human cardiomyocytes, raising the possibility that cell fusion may contribute to developing or maintaining polyploid cardiomyocytes in the human heart.
Keywords: Cardiomyocytes; Cell fusion; Copy number variation; Heart; Polyploid cardiomyocyte transcriptome and chromatin landscapes; Polyploidization; Somatic single nucleotide variants.
Conflict of interest statement
Competing interest declaration: The authors declare that they have no competing interests.
Figures
Similar articles
-
Polyploid cardiomyocytes: implications for heart regeneration.Development. 2021 Jul 15;148(14):dev199401. doi: 10.1242/dev.199401. Epub 2021 Jul 26. Development. 2021. PMID: 34897388 Free PMC article.
-
[Interconnection of parameters of the mitochondrial and myofibrillar apparatus of cardiomyocytes and ploidy and hypertrophy in certain mammalian species, differing in body mass].Tsitologiia. 1997;39(10):946-64. Tsitologiia. 1997. PMID: 9505342 Russian.
-
Myocardial Polyploidization Creates a Barrier to Heart Regeneration in Zebrafish.Dev Cell. 2018 Feb 26;44(4):433-446.e7. doi: 10.1016/j.devcel.2018.01.021. Dev Cell. 2018. PMID: 29486195 Free PMC article.
-
Advances in heart regeneration based on cardiomyocyte proliferation and regenerative potential of binucleated cardiomyocytes and polyploidization.Clin Sci (Lond). 2019 Jun 7;133(11):1229-1253. doi: 10.1042/CS20180560. Print 2019 Jun 14. Clin Sci (Lond). 2019. PMID: 31175264 Review.
-
Cardiomyocyte Polyploidy and Implications for Heart Regeneration.Annu Rev Physiol. 2020 Feb 10;82:45-61. doi: 10.1146/annurev-physiol-021119-034618. Epub 2019 Oct 4. Annu Rev Physiol. 2020. PMID: 31585517 Review.
References
-
- Swynghedauw B. Are adult cardiocytes still able to proliferate? Arch Mal Coeur Vaiss 96, 1225–1230 (2003). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
