

Phylogenetic position and plastid genome structure of Vietorchis, a mycoheterotrophic genus of Orchidaceae (subtribe Orchidinae) endemic to Vietnam
- PMID: 38855461
- PMCID: PMC11157612
- DOI: 10.3389/fpls.2024.1393225
Phylogenetic position and plastid genome structure of Vietorchis, a mycoheterotrophic genus of Orchidaceae (subtribe Orchidinae) endemic to Vietnam
Abstract
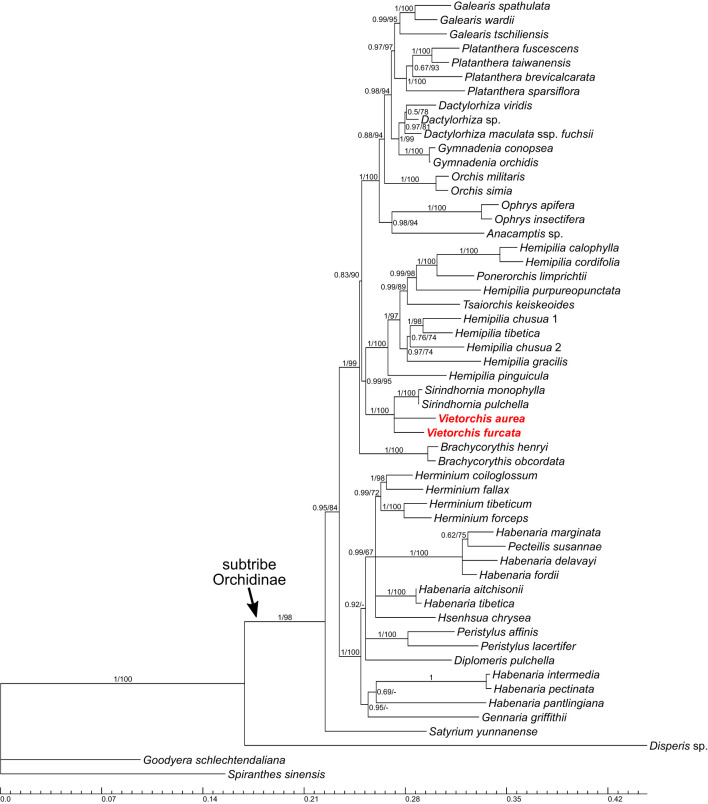
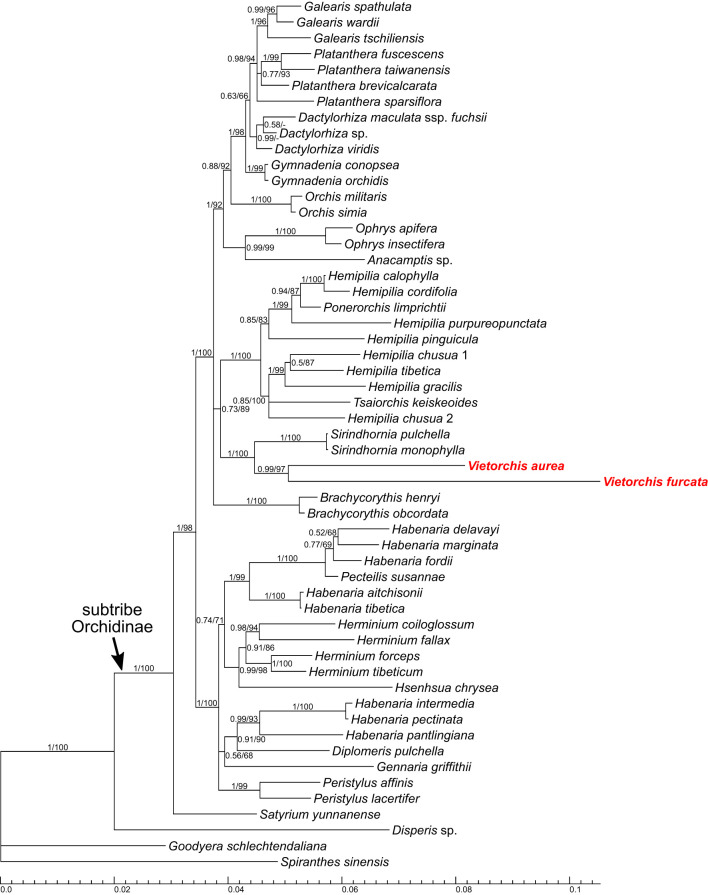
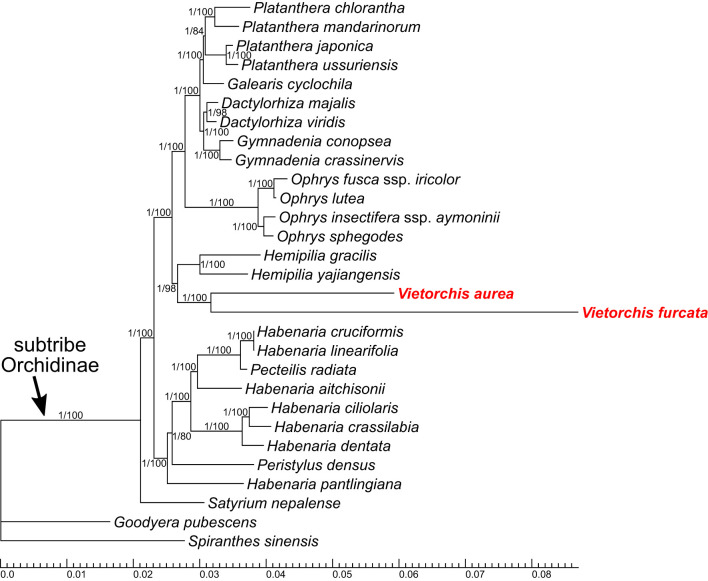
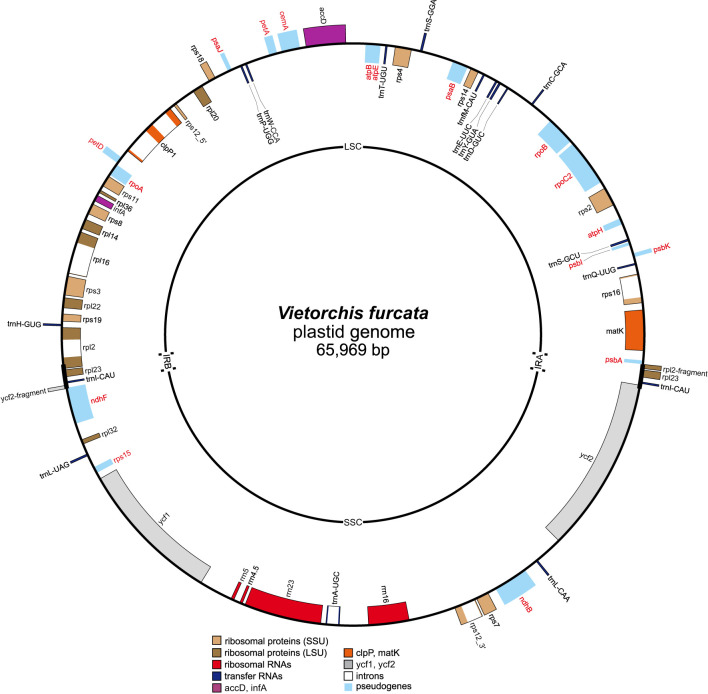
The orchid genus Vietorchis comprises three species, all discovered in the 21 century. Each of these species is achlorophyllous, mycoheterotrophic and is known to be endemic to Vietnam. The type species of the genus, V. aurea, occurs in a single location in northern Vietnam within a lowland limestone karstic area. Vietorchis furcata and V. proboscidea, in contrast, are confined to mountains of southern Vietnam, far away from any limestone formations. Taxonomic placement of Vietorchis remained uncertain for the reason of inconclusive morphological affinities. At the same time, the genus has never been included into molecular phylogenetic studies. We investigate the phylogenetic relationships of two species of Vietorchis (V. aurea and V. furcata) based on three DNA datasets: (1) a dataset comprising two nuclear regions, (2) a dataset comprising two plastid regions, and (3) a dataset employing data on the entire plastid genomes. Our phylogenetic reconstructions support the placement of Vietorchis into the subtribe Orchidinae (tribe Orchideae, subfamily Orchidoideae). This leads to a conclusion that the previously highlighted similarities in the rhizome morphology between Vietorchis and certain mycoheterotrophic genera of the subfamilies Epidendroideae and Vanilloideae are examples of a convergence. Vietorchis is deeply nested within Orchidinae, and therefore the subtribe Vietorchidinae is to be treated as a synonym of Orchidinae. In the obtained phylogenetic reconstructions, Vietorchis is sister to the photosynthetic genus Sirindhornia. Sirindhornia is restricted to limestone mountains, which allows to speculate that association with limestone karst is plesiomorphic for Vietorchis. Flower morphology is concordant with the molecular data in placing Vietorchis into Orchidinae and strongly supports the assignment of the genus to one of the two major clades within this subtribe. Within this clade, however, Vietorchis shows no close structural similarity with any of its genera; in particular, the proximity between Vietorchis and Sirindhornia has never been proposed. Finally, we assembled the plastid genome of V. furcata, which is 65969 bp long and contains 45 unique genes, being one of the most reduced plastomes in the subfamily Orchidoideae. The plastome of Vietorchis lacks any rearrangements in comparison with the closest studied autotrophic species, and possesses substantially contracted inverted repeats. No signs of positive selection acting on the protein-coding plastid sequences were detected.
Keywords: Silvorchis; Sirindhornia; genome reductive evolution; non-photosynthetic plants; taxonomy; tribe Orchideae.
Copyright © 2024 Samigullin, Logacheva, Averyanov, Zeng, Fu and Nuraliev.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


References
-
- Akaike H. (1974). A new look at the statistical model identification. IEEE Trans. Autom. Control 19, 716–723. doi: 10.1109/TAC.1974.1100705 - DOI
-
- Arias-Agudelo L. M., González F., Isaza J. P., Alzate J. F., Pabón-Mora N. (2019). Plastome reduction and gene content in New World Pilostyles (Apodanthaceae) unveils high similarities to African and Australian congeners. Mol. Phyl. Evol. 135, 193–202. doi: 10.1016/j.ympev.2019.03.014 - DOI - PubMed
-
- Averyanov L. V. (2008). The orchids of Vietnam. Illustrated survey. Part 1. Subfamilies Apostasioideae, Cypripedioideae and Spiranthoideae. Turczaninowia 11, 5–168.
-
- Averyanov L. V. (2010). The orchids of Vietnam. Illustrated survey. Part 2. Subfamily Orchidoideae. Turczaninowia 13, 5–98.
LinkOut - more resources
Full Text Sources