

Cell-autonomous role of leucine-rich repeat kinase in the protection of dopaminergic neuron survival
- PMID: 38856715
- PMCID: PMC11164531
- DOI: 10.7554/eLife.92673
Cell-autonomous role of leucine-rich repeat kinase in the protection of dopaminergic neuron survival
Abstract
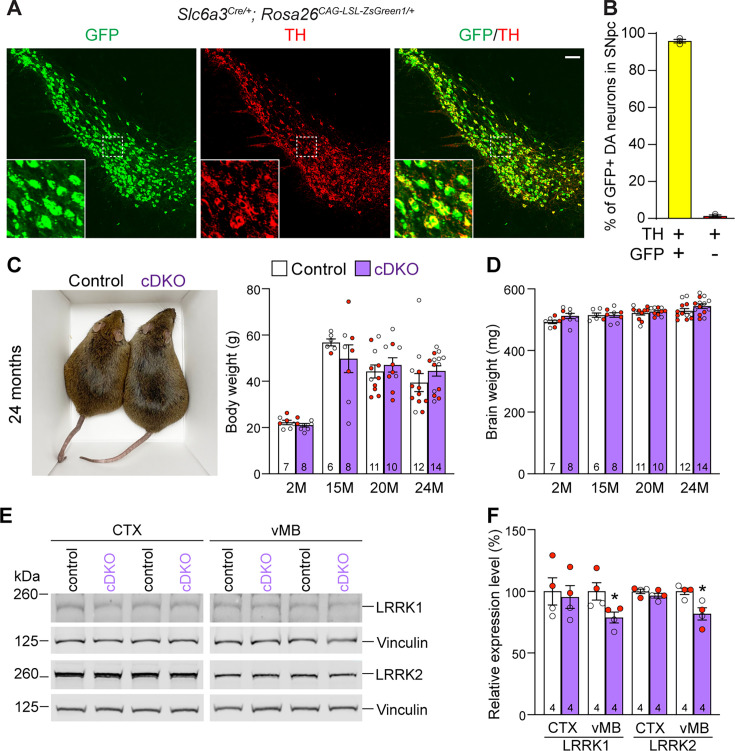
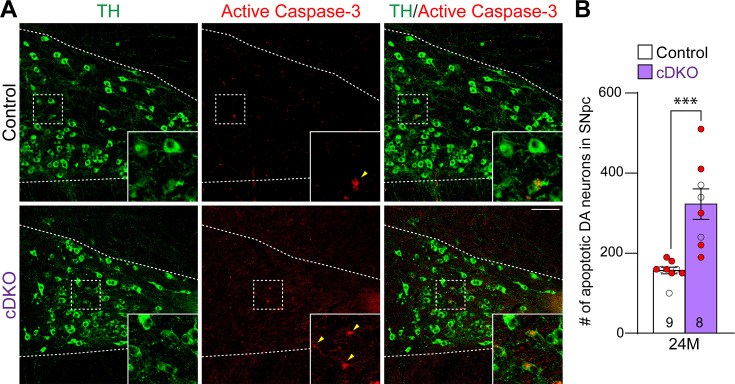
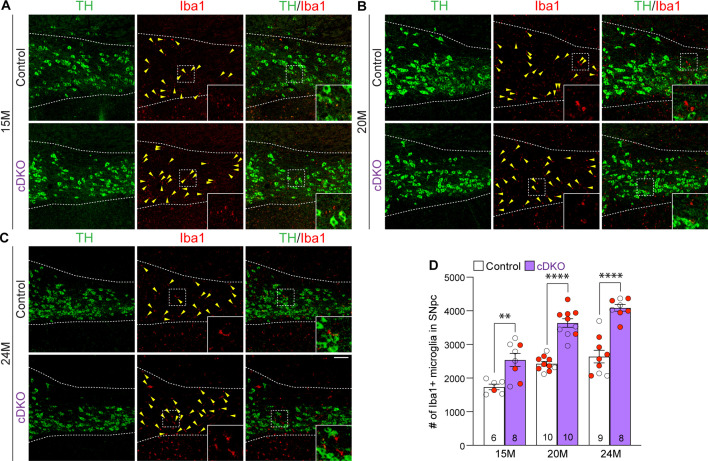
Mutations in leucine-rich repeat kinase 2 (LRRK2) are the most common genetic cause of Parkinson's disease (PD). However, whether LRRK2 mutations cause PD and degeneration of dopaminergic (DA) neurons via a toxic gain-of-function or a loss-of-function mechanism is unresolved and has pivotal implications for LRRK2-based PD therapies. In this study, we investigate whether Lrrk2 and its functional homolog Lrrk1 play a cell-intrinsic role in DA neuron survival through the development of DA neuron-specific Lrrk conditional double knockout (cDKO) mice. Unlike Lrrk germline DKO mice, DA neuron-restricted Lrrk cDKO mice exhibit normal mortality but develop age-dependent loss of DA neurons, as shown by the progressive reduction of DA neurons in the substantia nigra pars compacta (SNpc) at the ages of 20 and 24 months. Moreover, DA neurodegeneration is accompanied with increases in apoptosis and elevated microgliosis in the SNpc as well as decreases in DA terminals in the striatum, and is preceded by impaired motor coordination. Taken together, these findings provide the unequivocal evidence for the cell-intrinsic requirement of LRRK in DA neurons and raise the possibility that LRRK2 mutations may impair its protection of DA neurons, leading to DA neurodegeneration in PD.
Keywords: LRRK1; LRRK2; Parkinson's disease; conditional knock-out mice; dopaminergic neuron; mouse; neuroscience; substantia nigra pars compacta.
© 2023, Kang, Huang et al.
Conflict of interest statement
JK, GH, LM, YT, AS, PC, JS No competing interests declared
Figures
















Update of
-
Cell autonomous role of leucine-rich repeat kinase in protection of dopaminergic neuron survival.bioRxiv [Preprint]. 2024 Mar 7:2023.10.06.561293. doi: 10.1101/2023.10.06.561293. bioRxiv. 2024. Update in: Elife. 2024 Jun 10;12:RP92673. doi: 10.7554/eLife.92673. PMID: 37873418 Free PMC article. Updated. Preprint.
References
-
- An X-K, Peng R, Li T, Burgunder J-M, Wu Y, Chen W-J, Zhang J-H, Wang Y-C, Xu Y-M, Gou Y-R, Yuan G-G, Zhang Z-J. LRRK2 Gly2385Arg variant is a risk factor of Parkinson’s disease among Han-Chinese from mainland China. European Journal of Neurology. 2008;15:301–305. doi: 10.1111/j.1468-1331.2007.02052.x. - DOI - PubMed
-
- Blauwendraat C, Reed X, Kia DA, Gan-Or Z, Lesage S, Pihlstrøm L, Guerreiro R, Gibbs JR, Sabir M, Ahmed S, Ding J, Alcalay RN, Hassin-Baer S, Pittman AM, Brooks J, Edsall C, Hernandez DG, Chung SJ, Goldwurm S, Toft M, Schulte C, Bras J, Wood NW, Brice A, Morris HR, Scholz SW, Nalls MA, Singleton AB, Cookson MR, COURAGE-PD (Comprehensive Unbiased Risk Factor Assessment for Genetics and Environment in Parkinson’s Disease) Consortium, the French Parkinson’s Disease Consortium, and the International Parkinson’s Disease Genomics Consortium (IPDGC) Frequency of loss of function variants in LRRK2 in Parkinson disease. JAMA Neurology. 2018;75:1416–1422. doi: 10.1001/jamaneurol.2018.1885. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
