

The role of lysosomal phospholipase A2 in the catabolism of bis(monoacylglycerol)phosphate and association with phospholipidosis
- PMID: 38857781
- PMCID: PMC11277439
- DOI: 10.1016/j.jlr.2024.100574
The role of lysosomal phospholipase A2 in the catabolism of bis(monoacylglycerol)phosphate and association with phospholipidosis
Abstract
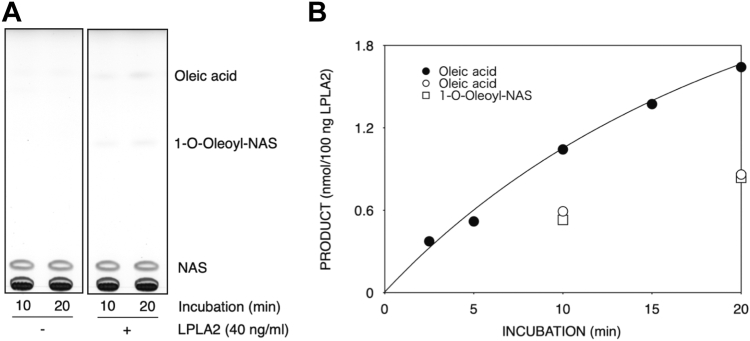
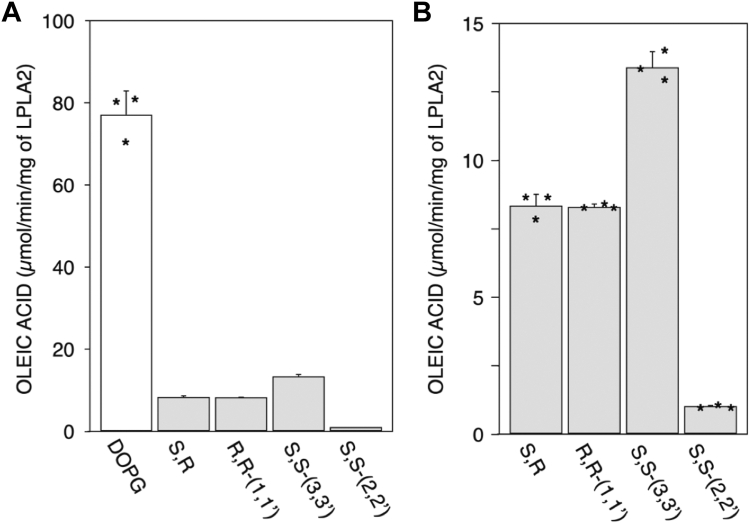
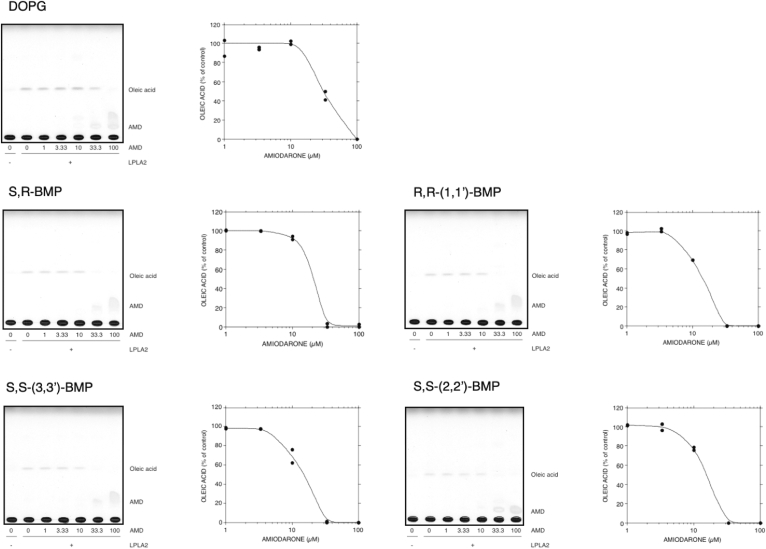
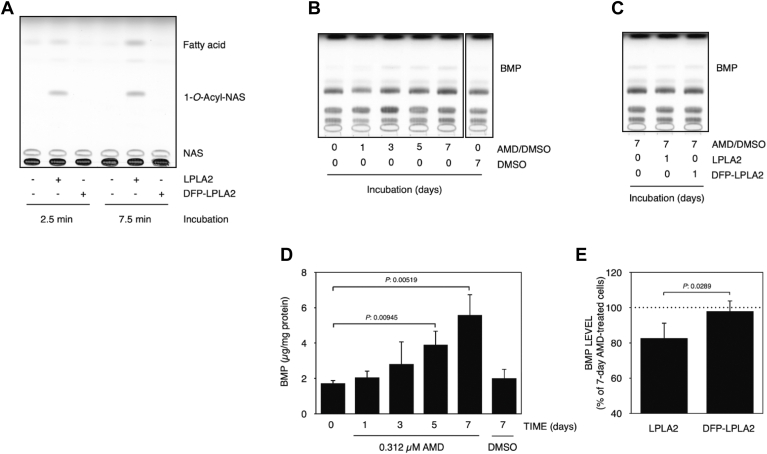
Bis(monoacylglycerol)phosphate (BMP) is an acidic glycerophospholipid localized to late endosomes and lysosomes. However, the metabolism of BMP is poorly understood. Because many drugs that cause phospholipidosis inhibit lysosomal phospholipase A2 (LPLA2, PLA2G15, LYPLA3) activity, we investigated whether this enzyme has a role in BMPcatabolism. The incubation of recombinant human LPLA2 (hLPLA2) and liposomes containing the naturally occurring BMP (sn-(2-oleoyl-3-hydroxy)-glycerol-1-phospho-sn-1'-(2'-oleoyl-3'-hydroxy)-glycerol (S,S-(2,2',C18:1)-BMP) resulted in the deacylation of this BMP isomer. The deacylation rate was 70 times lower than that of dioleoyl phosphatidylglycerol (DOPG), an isomer and precursor of BMP. The release rates of oleic acid from DOPG and four BMP stereoisomers by LPLA2 differed. The rank order of the rates of hydrolysis were DOPG>S,S-(3,3',C18:1)-BMP>R,S-(3,1',C18:1)-BMP>R,R-(1,1',C18:1)>S,S-(2,2')-BMP. The cationic amphiphilic drug amiodarone (AMD) inhibited the deacylation of DOPG and BMP isomers by hLPLA2 in a concentration-dependent manner. Under these experimental conditions, the IC50s of amiodarone-induced inhibition of the four BMP isomers and DOPG were less than 20 μM and approximately 30 μM, respectively. BMP accumulation was observed in AMD-treated RAW 264.7 cells. The accumulated BMP was significantly reduced by exogenous treatment of cells with active recombinant hLPLA2 but not with diisopropylfluorophosphate-inactivated recombinant hLPLA2. Finally, a series of cationic amphiphilic drugs known to cause phospholipidosis were screened for inhibition of LPLA2 activity as measured by either the transacylation or fatty acid hydrolysis of BMP or phosphatidylcholine as substrates. Fifteen compounds demonstrated significant inhibition with IC50s ranging from 6.8 to 63.3 μM. These results indicate that LPLA2 degrades BMP isomers with different substrate specificities under acidic conditions and may be the key enzyme associated with BMP accumulation in drug-induced phospholipidosis.
Keywords: amiodarone; bis(monoacylglycerol)phosphate; cationic amphiphilic drug; lysosomal phospholipase; phospholipase A2 group 15; phospholipidosis.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interests The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. The author is an Editorial Board Member/Editor-in-Chief/Associate Editor/Guest Editor for The Journal of Lipid Research and was not involved in the editorial review or the decision to publish this article.
Figures




Similar articles
-
Isomerization of bis(monoacylglycero)phosphate by acyl migration.J Lipid Res. 2025 May;66(5):100789. doi: 10.1016/j.jlr.2025.100789. Epub 2025 Mar 29. J Lipid Res. 2025. PMID: 40164336 Free PMC article.
-
Inhibition of lysosomal phospholipase A2 predicts drug-induced phospholipidosis.J Lipid Res. 2021;62:100089. doi: 10.1016/j.jlr.2021.100089. Epub 2021 Jun 1. J Lipid Res. 2021. PMID: 34087196 Free PMC article. Review.
-
Preferential hydrolysis of truncated oxidized glycerophospholipids by lysosomal phospholipase A2.J Lipid Res. 2017 Feb;58(2):339-349. doi: 10.1194/jlr.M070730. Epub 2016 Dec 19. J Lipid Res. 2017. PMID: 27993948 Free PMC article.
-
Di-22:6-bis(monoacylglycerol)phosphate: A clinical biomarker of drug-induced phospholipidosis for drug development and safety assessment.Toxicol Appl Pharmacol. 2014 Sep 15;279(3):467-476. doi: 10.1016/j.taap.2014.06.014. Epub 2014 Jun 23. Toxicol Appl Pharmacol. 2014. PMID: 24967688
-
Lysosomal phospholipase A2.Biochim Biophys Acta Mol Cell Biol Lipids. 2019 Jun;1864(6):932-940. doi: 10.1016/j.bbalip.2018.07.012. Epub 2018 Aug 2. Biochim Biophys Acta Mol Cell Biol Lipids. 2019. PMID: 30077006 Free PMC article. Review.
Cited by
-
Hepatic lipid remodeling in cold exposure uncovers direct regulation of bis(monoacylglycero)phosphate lipids by phospholipase A2 group XV.Cell Metab. 2025 Jun 3;37(6):1413-1425.e6. doi: 10.1016/j.cmet.2025.04.015. Epub 2025 May 14. Cell Metab. 2025. PMID: 40373767
-
Molecular determinants of phospholipid treatment to reduce intracellular cholesterol accumulation in NPC1 deficiency.J Biol Chem. 2024 Nov;300(11):107889. doi: 10.1016/j.jbc.2024.107889. Epub 2024 Oct 11. J Biol Chem. 2024. PMID: 39395801 Free PMC article.
-
SPNS1 variants cause multiorgan disease and implicate lysophospholipid transport as critical for mTOR-regulated lipid homeostasis.J Clin Invest. 2025 Jul 3;135(17):e193099. doi: 10.1172/JCI193099. eCollection 2025 Sep 2. J Clin Invest. 2025. PMID: 40608416 Free PMC article.
-
PLA2G15 is a BMP hydrolase and its targeting ameliorates lysosomal disease.Nature. 2025 Jun;642(8067):474-483. doi: 10.1038/s41586-025-08942-y. Epub 2025 May 7. Nature. 2025. PMID: 40335701 Free PMC article.
-
Isomerization of bis(monoacylglycero)phosphate by acyl migration.J Lipid Res. 2025 May;66(5):100789. doi: 10.1016/j.jlr.2025.100789. Epub 2025 Mar 29. J Lipid Res. 2025. PMID: 40164336 Free PMC article.
References
-
- Amidon B., Schmitt J.D., Thuren T., King L., Waite M. Biosynthetic conversion of phosphatidylglycerol to sn-1:sn-1' bis(monoacylglycerol) phosphate in a macrophage-like cell line. Biochemistry. 1995;34:5554–5560. - PubMed
-
- Amidon B., Brown A., Waite M. Transacylase and phospholipases in the synthesis of bis(monoacylglycero)phosphate. Biochemistry. 1996;35:13995–14002. - PubMed
-
- Shinozaki K., Waite M. A novel phosphatidylglycerol-selective phospholipase A2 from macrophages. Biochemistry. 1999;38:1669–1675. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Miscellaneous
