

Sleep Disruption Precedes Forebrain Synaptic Tau Burden and Contributes to Cognitive Decline in a Sex-Dependent Manner in the P301S Tau Transgenic Mouse Model
- PMID: 38858068
- PMCID: PMC11209651
- DOI: 10.1523/ENEURO.0004-24.2024
Sleep Disruption Precedes Forebrain Synaptic Tau Burden and Contributes to Cognitive Decline in a Sex-Dependent Manner in the P301S Tau Transgenic Mouse Model
Abstract
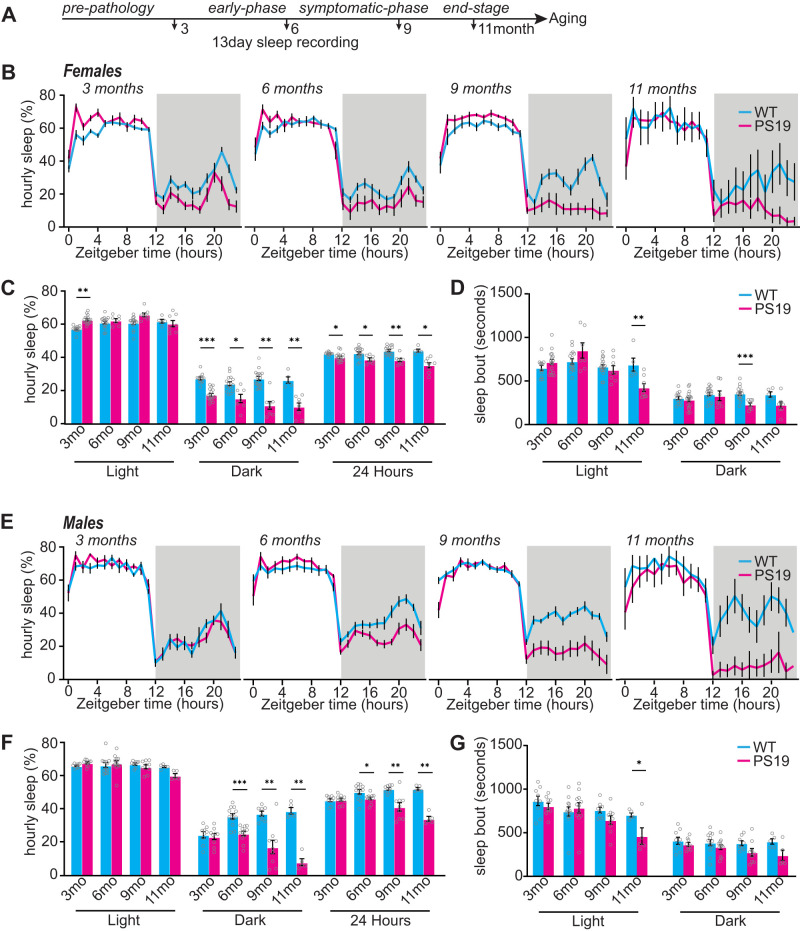
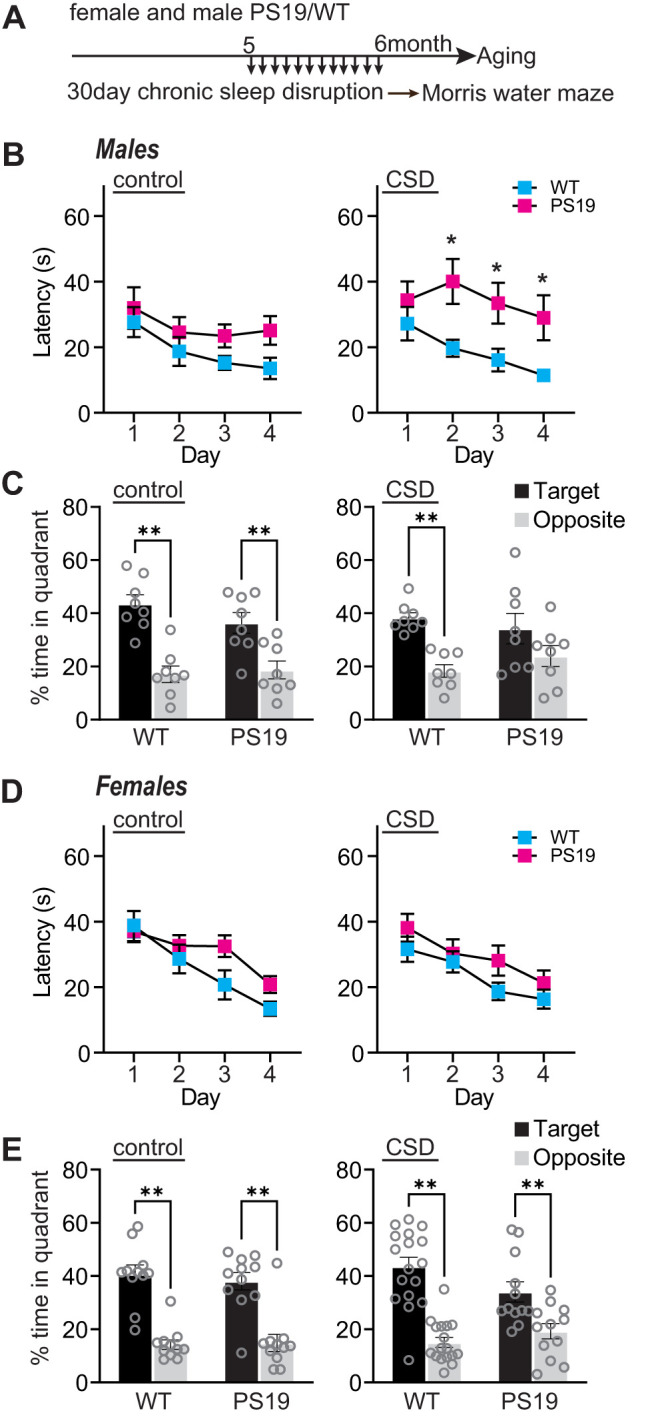
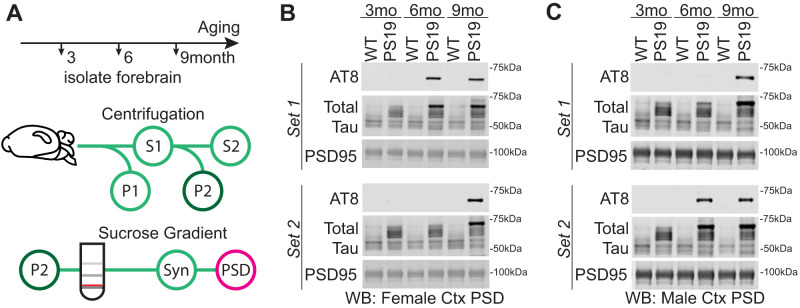
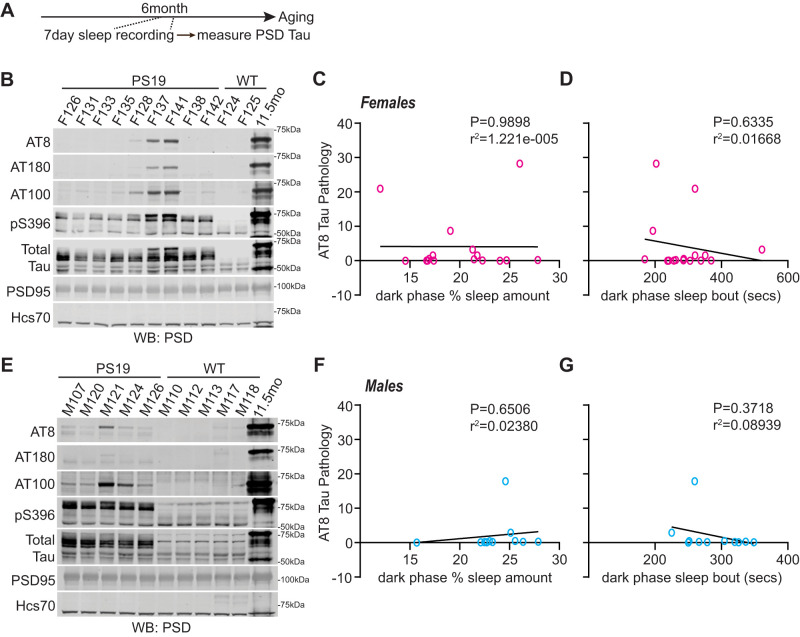
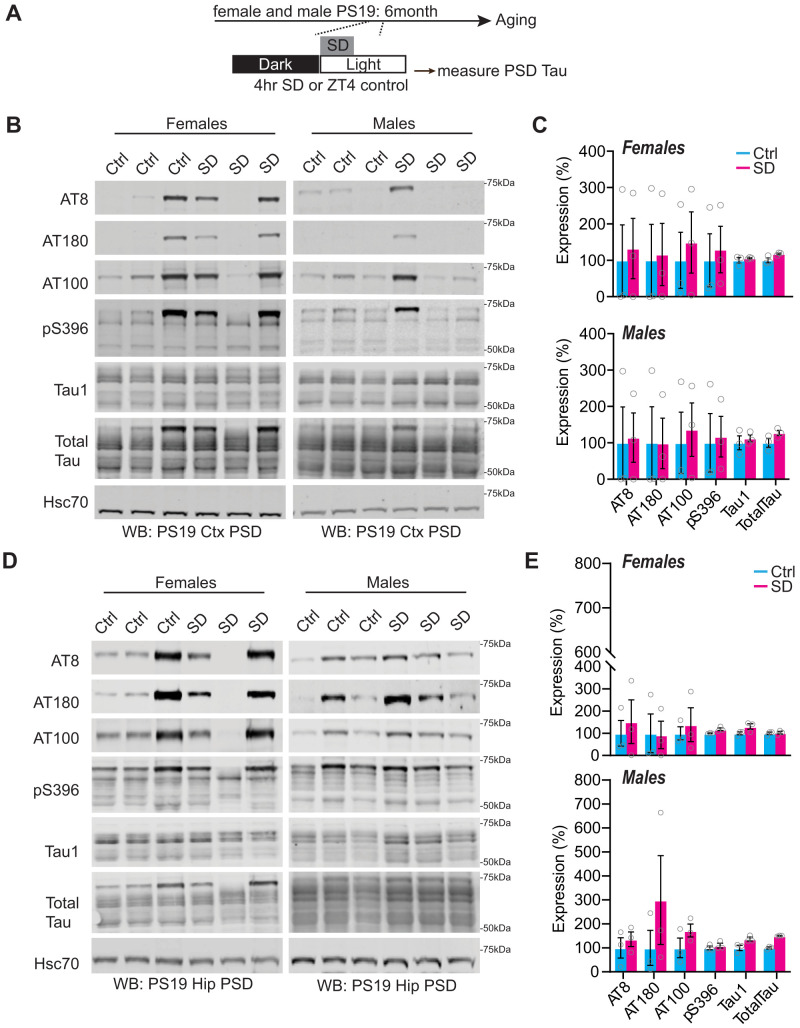
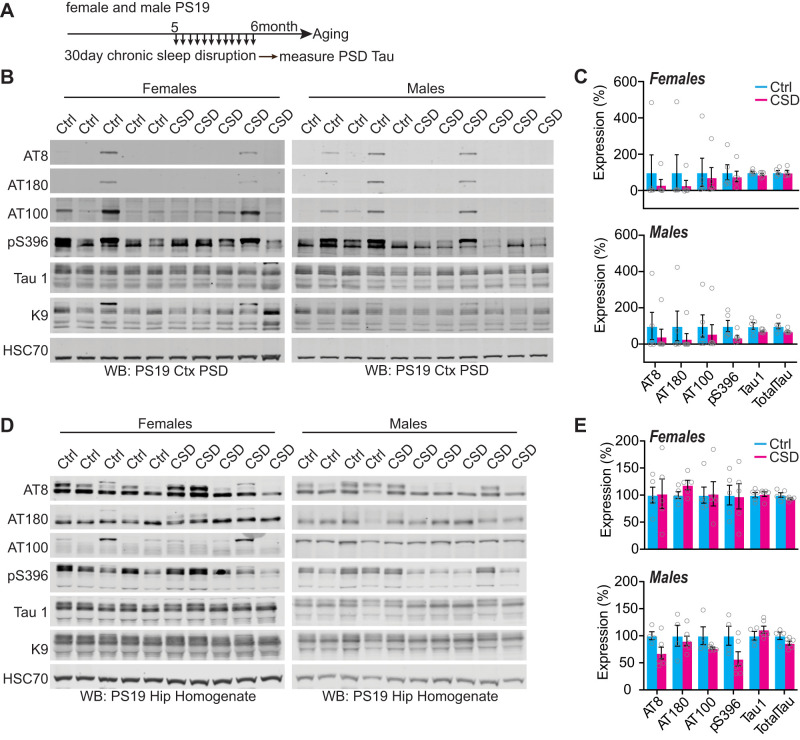
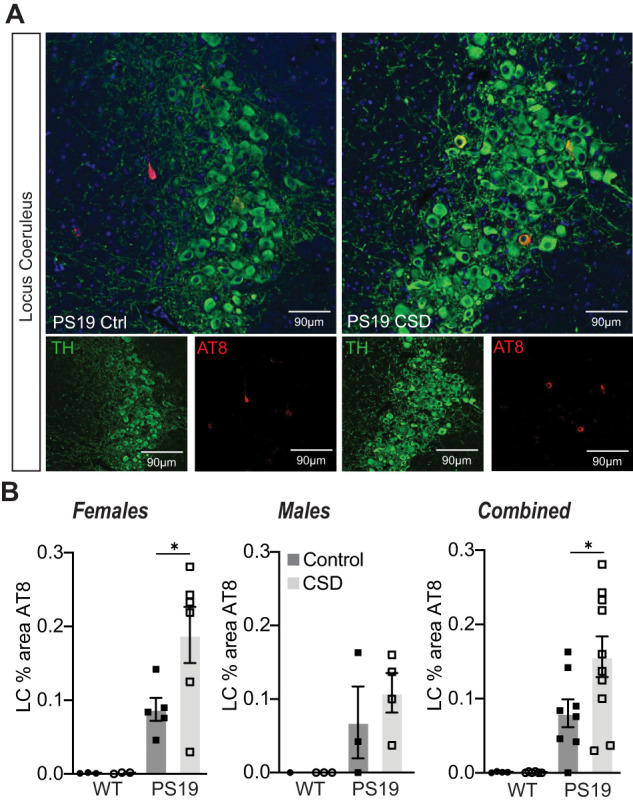
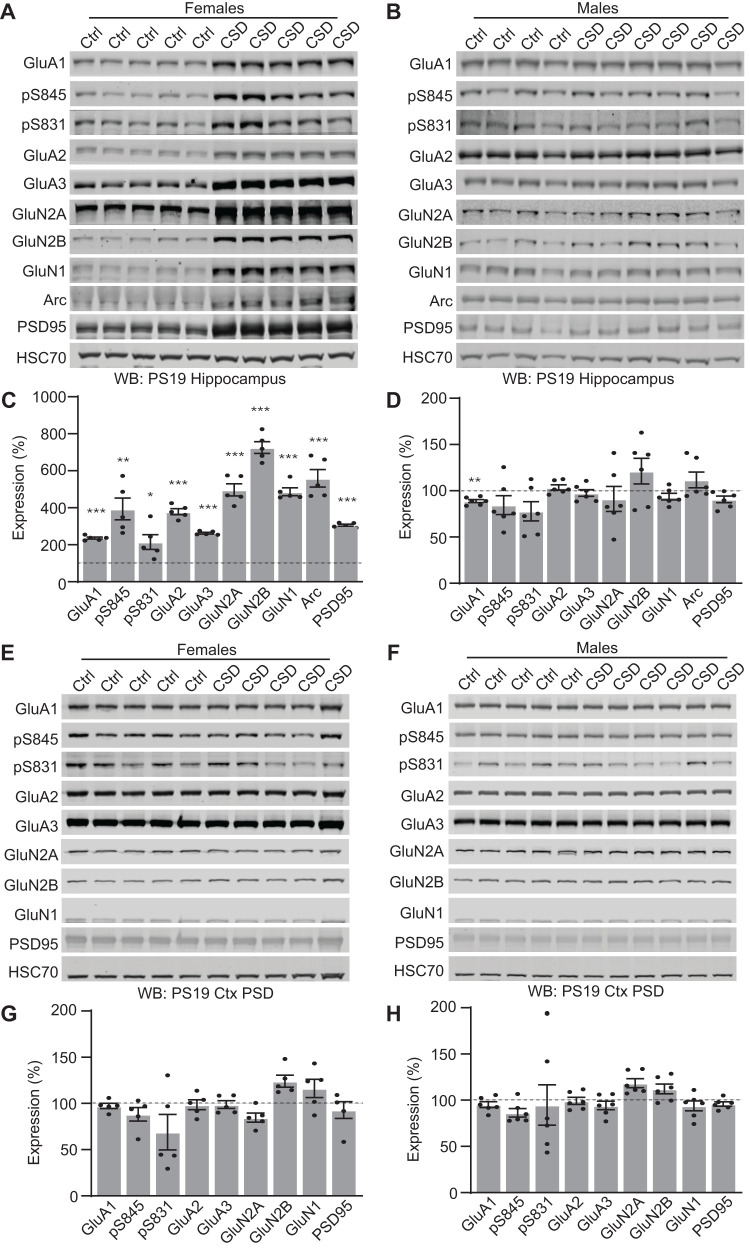
Sleep disruption and impaired synaptic processes are common features in neurodegenerative diseases, including Alzheimer's disease (AD). Hyperphosphorylated Tau is known to accumulate at neuronal synapses in AD, contributing to synapse dysfunction. However, it remains unclear how sleep disruption and synapse pathology interact to contribute to cognitive decline. Here, we examined sex-specific onset and consequences of sleep loss in AD/tauopathy model PS19 mice. Using a piezoelectric home-cage monitoring system, we showed PS19 mice exhibited early-onset and progressive hyperarousal, a selective dark-phase sleep disruption, apparent at 3 months in females and 6 months in males. Using the Morris water maze test, we report that chronic sleep disruption (CSD) accelerated the onset of decline of hippocampal spatial memory in PS19 males only. Hyperarousal occurs well in advance of robust forebrain synaptic Tau burden that becomes apparent at 6-9 months. To determine whether a causal link exists between sleep disruption and synaptic Tau hyperphosphorylation, we examined the correlation between sleep behavior and synaptic Tau, or exposed mice to acute or chronic sleep disruption at 6 months. While we confirm that sleep disruption is a driver of Tau hyperphosphorylation in neurons of the locus ceruleus, we were unable to show any causal link between sleep loss and Tau burden in forebrain synapses. Despite the finding that hyperarousal appears earlier in females, female cognition was resilient to the effects of sleep disruption. We conclude sleep disruption interacts with the synaptic Tau burden to accelerate the onset of cognitive decline with greater vulnerability in males.
Keywords: Alzheimer's disease; Tau; biological sex; sleep; sleep disruption; synapse.
Copyright © 2024 Martin et al.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Update of
-
Sleep disruption precedes forebrain synaptic Tau burden and contributes to cognitive decline in a sex-dependent manner in the P301S Tau transgenic mouse model.bioRxiv [Preprint]. 2023 Jun 9:2023.06.07.544101. doi: 10.1101/2023.06.07.544101. bioRxiv. 2023. Update in: eNeuro. 2024 Jun 26;11(6):ENEURO.0004-24.2024. doi: 10.1523/ENEURO.0004-24.2024. PMID: 37333395 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases