

A folding motif formed with an expanded genetic alphabet
- PMID: 38858518
- PMCID: PMC11446821
- DOI: 10.1038/s41557-024-01552-7
A folding motif formed with an expanded genetic alphabet
Abstract
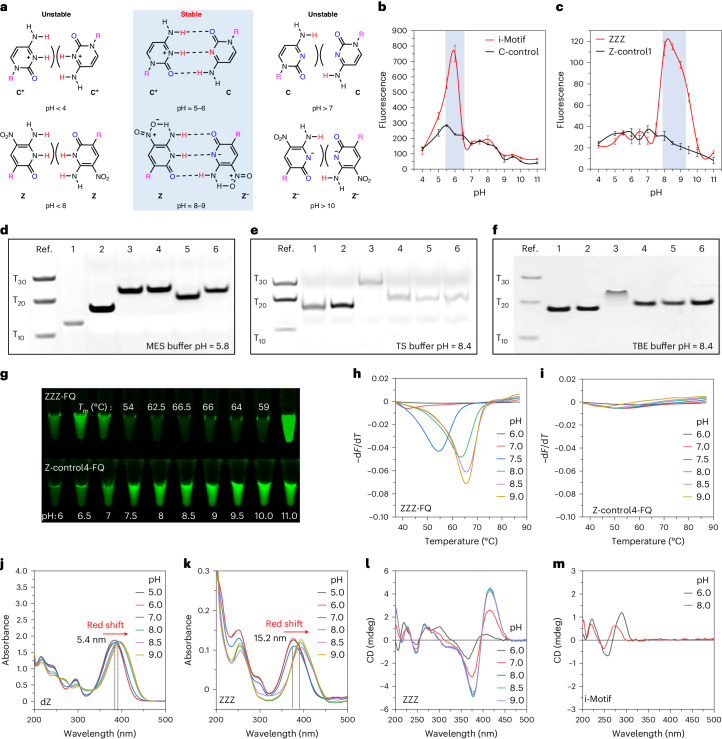
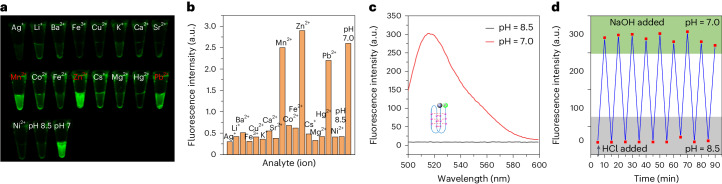
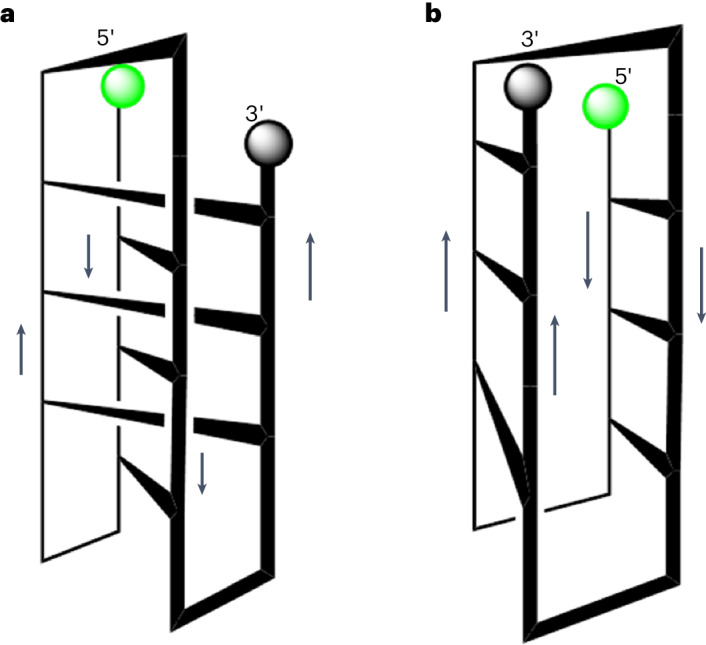
Adding synthetic nucleotides to DNA increases the linear information density of DNA molecules. Here we report that it also can increase the diversity of their three-dimensional folds. Specifically, an additional nucleotide (dZ, with a 5-nitro-6-aminopyridone nucleobase), placed at twelve sites in a 23-nucleotides-long DNA strand, creates a fairly stable unimolecular structure (that is, the folded Z-motif, or fZ-motif) that melts at 66.5 °C at pH 8.5. Spectroscopic, gel and two-dimensional NMR analyses show that the folded Z-motif is held together by six reverse skinny dZ-:dZ base pairs, analogous to the crystal structure of the free heterocycle. Fluorescence tagging shows that the dZ-:dZ pairs join parallel strands in a four-stranded compact down-up-down-up fold. These have two possible structures: one with intercalated dZ-:dZ base pairs, the second without intercalation. The intercalated structure would resemble the i-motif formed by dC:dC+-reversed pairing at pH ≤ 6.5. This fZ-motif may therefore help DNA form compact structures needed for binding and catalysis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Conner, B. N., Takano, T., Tanaka, S., Itakura, K. & Dickerson, R. E. The molecular structure of d(ICpCpGpG), a fragment of right-handed double helical A-DNA. Nature295, 294–299 (1982). - PubMed
-
- Panayotatos, N. & Wells, R. D. Cruciform structures in supercoiled DNA. Nature289, 466–470 (1981). - PubMed
MeSH terms
Substances
Grants and funding
- S10 OD028753/OD/NIH HHS/United States
- 1R01GM128186/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- 21877031/National Natural Science Foundation of China (National Science Foundation of China)
- R01 GM141391/GM/NIGMS NIH HHS/United States
- R01 GM128186/GM/NIGMS NIH HHS/United States
- 1R01GM141391-01A1/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- 21991080/National Natural Science Foundation of China (National Science Foundation of China)
- 22274044/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
