

K128 ubiquitination constrains RAS activity by expanding its binding interface with GAP proteins
- PMID: 38858602
- PMCID: PMC11251195
- DOI: 10.1038/s44318-024-00146-w
K128 ubiquitination constrains RAS activity by expanding its binding interface with GAP proteins
Abstract
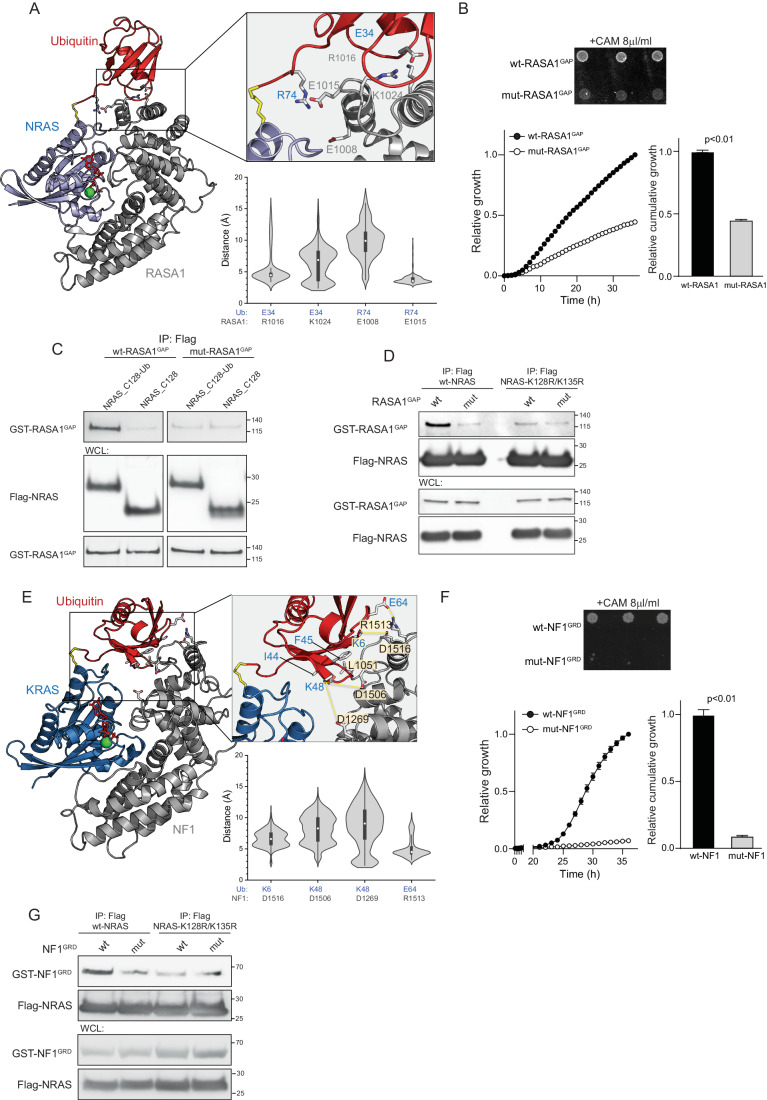
The RAS pathway is among the most frequently activated signaling nodes in cancer. However, the mechanisms that alter RAS activity in human pathologies are not entirely understood. The most prevalent post-translational modification within the GTPase core domain of NRAS and KRAS is ubiquitination at lysine 128 (K128), which is significantly decreased in cancer samples compared to normal tissue. Here, we found that K128 ubiquitination creates an additional binding interface for RAS GTPase-activating proteins (GAPs), NF1 and RASA1, thus increasing RAS binding to GAP proteins and promoting GAP-mediated GTP hydrolysis. Stimulation of cultured cancer cells with growth factors or cytokines transiently induces K128 ubiquitination and restricts the extent of wild-type RAS activation in a GAP-dependent manner. In KRAS mutant cells, K128 ubiquitination limits tumor growth by restricting RAL/ TBK1 signaling and negatively regulating the autocrine circuit induced by mutant KRAS. Reduction of K128 ubiquitination activates both wild-type and mutant RAS signaling and elicits a senescence-associated secretory phenotype, promoting RAS-driven pancreatic tumorigenesis.
Keywords: NF1; RAS Interactome; RAS Signaling; Senescence-Associated Secretory Phenotype; Ubiquitination.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Peptides containing a consensus Ras binding sequence from Raf-1 and theGTPase activating protein NF1 inhibit Ras function.Proc Natl Acad Sci U S A. 1996 Feb 20;93(4):1577-81. doi: 10.1073/pnas.93.4.1577. Proc Natl Acad Sci U S A. 1996. PMID: 8643674 Free PMC article.
-
Neurofibromin GTPase-activating protein-related domains restore normal growth in Nf1-/- cells.J Biol Chem. 2001 Mar 9;276(10):7240-5. doi: 10.1074/jbc.M009202200. Epub 2000 Nov 15. J Biol Chem. 2001. PMID: 11080503
-
MiR-21/RASA1 axis affects malignancy of colon cancer cells via RAS pathways.World J Gastroenterol. 2015 Feb 7;21(5):1488-97. doi: 10.3748/wjg.v21.i5.1488. World J Gastroenterol. 2015. PMID: 25663768 Free PMC article.
-
Calmodulin and IQGAP1 activation of PI3Kα and Akt in KRAS, HRAS and NRAS-driven cancers.Biochim Biophys Acta Mol Basis Dis. 2018 Jun;1864(6 Pt B):2304-2314. doi: 10.1016/j.bbadis.2017.10.032. Epub 2017 Oct 31. Biochim Biophys Acta Mol Basis Dis. 2018. PMID: 29097261 Review.
-
After 95 years, it's time to eRASe JMML.Blood Rev. 2020 Sep;43:100652. doi: 10.1016/j.blre.2020.100652. Epub 2020 Jan 16. Blood Rev. 2020. PMID: 31980238 Review.
Cited by
-
The differential interactomes of the KRAS splice variants identify BIRC6 as a ubiquitin ligase for KRAS4A.Cell Rep. 2025 Jan 28;44(1):115087. doi: 10.1016/j.celrep.2024.115087. Epub 2024 Dec 19. Cell Rep. 2025. PMID: 39705142 Free PMC article.
-
USP25 maintains KRAS expression and inhibiting the deubiquitinase suppresses KRAS signaling in human cancer.J Biol Chem. 2025 Jul;301(7):110337. doi: 10.1016/j.jbc.2025.110337. Epub 2025 Jun 3. J Biol Chem. 2025. PMID: 40473213 Free PMC article.
-
Pioneer in Molecular Biology: Conformational Ensembles in Molecular Recognition, Allostery, and Cell Function.J Mol Biol. 2025 Jun 1;437(11):169044. doi: 10.1016/j.jmb.2025.169044. Epub 2025 Feb 25. J Mol Biol. 2025. PMID: 40015370 Review.
-
Molecular principles underlying aggressive cancers.Signal Transduct Target Ther. 2025 Feb 17;10(1):42. doi: 10.1038/s41392-025-02129-7. Signal Transduct Target Ther. 2025. PMID: 39956859 Free PMC article. Review.
-
Computational studies on the functional and structural impact of pathogenic mutations in enzymes.Protein Sci. 2025 Apr;34(4):e70081. doi: 10.1002/pro.70081. Protein Sci. 2025. PMID: 40116283 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
