

Haplotype-resolved T2T genome assemblies and pangenome graph of pear reveal diverse patterns of allele-specific expression and the genomic basis of fruit quality traits
- PMID: 38859586
- PMCID: PMC11574287
- DOI: 10.1016/j.xplc.2024.101000
Haplotype-resolved T2T genome assemblies and pangenome graph of pear reveal diverse patterns of allele-specific expression and the genomic basis of fruit quality traits
Abstract
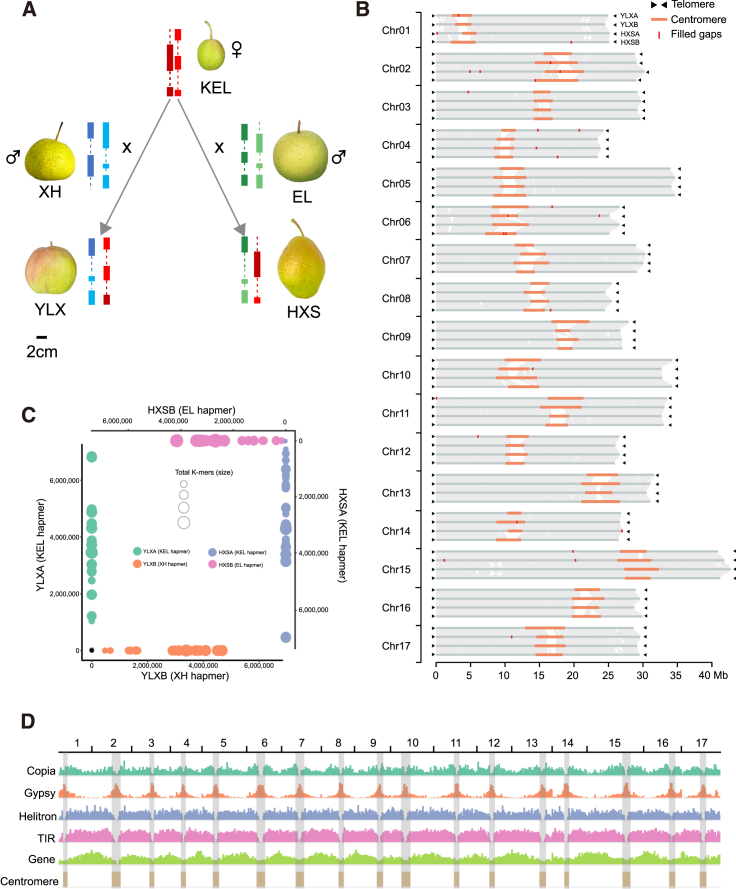
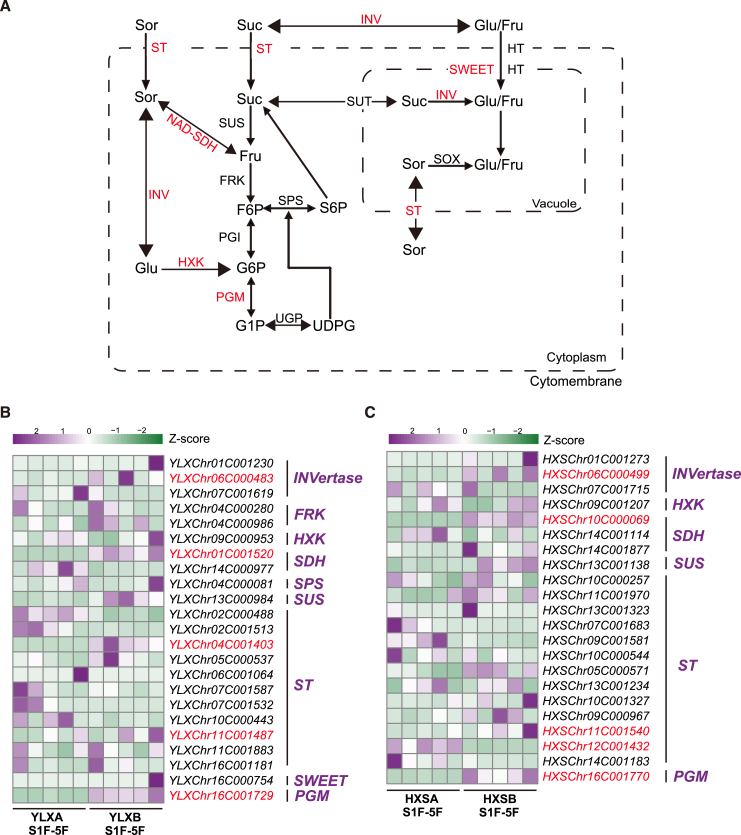
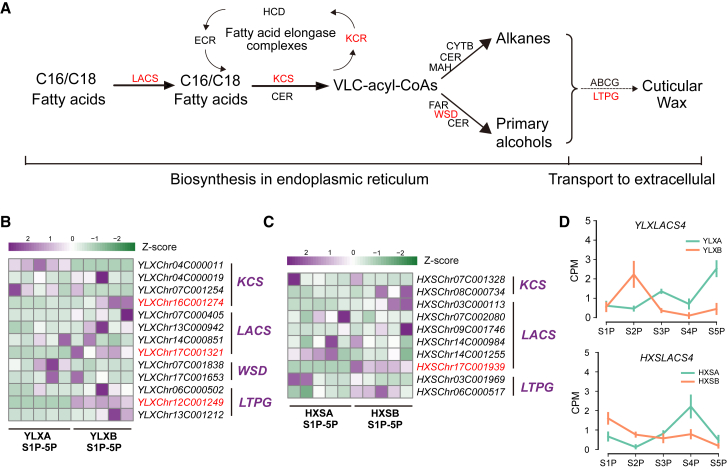
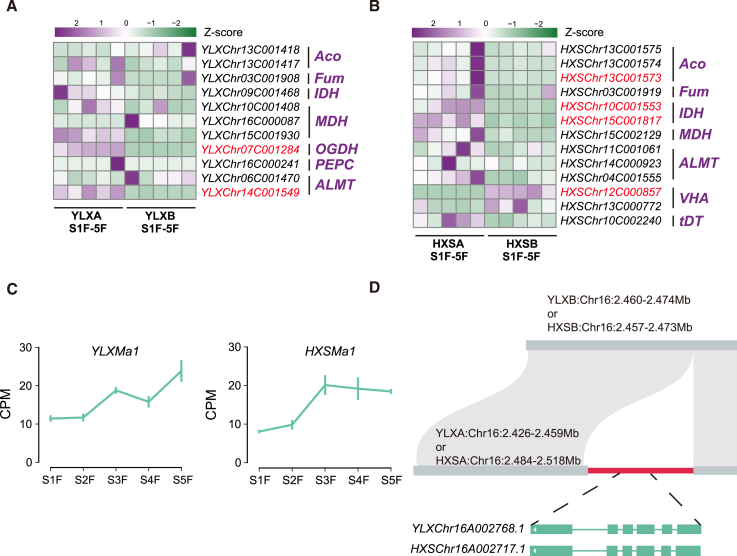
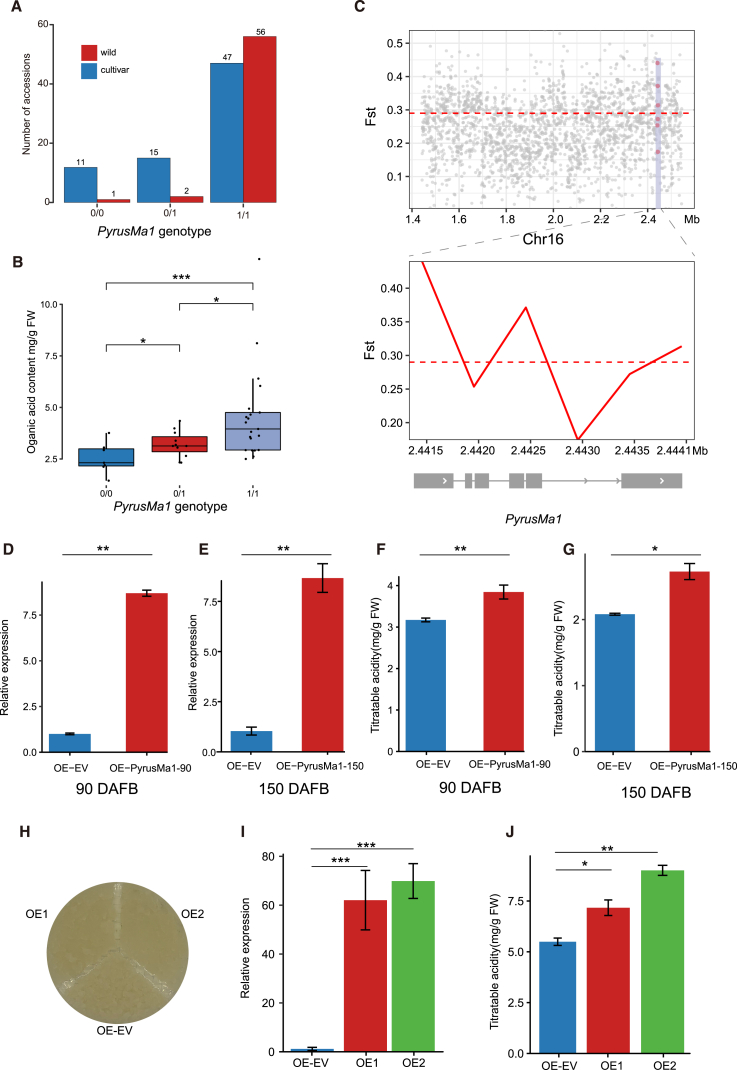
Hybrid crops often exhibit increased yield and greater resilience, yet the genomic mechanism(s) underlying hybrid vigor or heterosis remain unclear, hindering our ability to predict the expression of phenotypic traits in hybrid breeding. Here, we generated haplotype-resolved T2T genome assemblies of two pear hybrid varieties, 'Yuluxiang' (YLX) and 'Hongxiangsu' (HXS), which share the same maternal parent but differ in their paternal parents. We then used these assemblies to explore the genome-scale landscape of allele-specific expression (ASE) and create a pangenome graph for pear. ASE was observed for close to 6000 genes in both hybrid cultivars. A subset of ASE genes related to aspects of fruit quality such as sugars, organic acids, and cuticular wax were identified, suggesting their important contributions to heterosis. Specifically, Ma1, a gene regulating fruit acidity, is absent in the paternal haplotypes of HXS and YLX. A pangenome graph was built based on our assemblies and seven published pear genomes. Resequencing data for 139 cultivated pear genotypes (including 97 genotypes sequenced here) were subsequently aligned to the pangenome graph, revealing numerous structural variant hotspots and selective sweeps during pear diversification. As predicted, the Ma1 allele was found to be absent in varieties with low organic acid content, and this association was functionally validated by Ma1 overexpression in pear fruit and calli. Overall, these results reveal the contributions of ASE to fruit-quality heterosis and provide a robust pangenome reference for high-resolution allele discovery and association mapping.
Keywords: allele-specific expression; fruit quality traits; haplotype-resolved assembly; pangenome graph; pear.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Transcriptomic and Gas Chromatography-Mass Spectrometry Metabolomic Profiling Analysis of the Epidermis Provides Insights into Cuticular Wax Regulation in Developing 'Yuluxiang' Pear Fruit.J Agric Food Chem. 2019 Jul 31;67(30):8319-8331. doi: 10.1021/acs.jafc.9b01899. Epub 2019 Jul 19. J Agric Food Chem. 2019. PMID: 31287308
-
Haplotype-resolved, gap-free genome assemblies provide insights into the divergence between Asian and European pears.Nat Genet. 2025 Aug;57(8):2040-2051. doi: 10.1038/s41588-025-02273-4. Epub 2025 Aug 6. Nat Genet. 2025. PMID: 40770574 Free PMC article.
-
Transcriptome and Resequencing Analyses Provide Insight into Differences in Organic Acid Accumulation in Two Pear Varieties.Int J Mol Sci. 2021 Sep 6;22(17):9622. doi: 10.3390/ijms22179622. Int J Mol Sci. 2021. PMID: 34502530 Free PMC article.
-
A survey of sequence-to-graph mapping algorithms in the pangenome era.Genome Biol. 2025 May 22;26(1):138. doi: 10.1186/s13059-025-03606-6. Genome Biol. 2025. PMID: 40405275 Free PMC article. Review.
-
Computational Strategies for Eukaryotic Pangenome Analyses.2020 May 1. In: Tettelin H, Medini D, editors. The Pangenome: Diversity, Dynamics and Evolution of Genomes [Internet]. Cham (CH): Springer; 2020. 2020 May 1. In: Tettelin H, Medini D, editors. The Pangenome: Diversity, Dynamics and Evolution of Genomes [Internet]. Cham (CH): Springer; 2020. PMID: 32633910 Free Books & Documents. Review.
Cited by
-
Graph pangenome reveals the regulation of malate content in blood-fleshed peach by NAC transcription factors.Genome Biol. 2025 Jan 9;26(1):7. doi: 10.1186/s13059-024-03470-w. Genome Biol. 2025. PMID: 39789611 Free PMC article.
-
The telomere-to-telomere genome of Pucai () (Typha angustifolia L.): a distinctive semiaquatic vegetable with lignin and chlorophyll as quality characteristics.Hortic Res. 2025 Mar 11;12(7):uhaf079. doi: 10.1093/hr/uhaf079. eCollection 2025 Jul. Hortic Res. 2025. PMID: 40343350 Free PMC article.
-
Haplotype-resolved T2T genome assembly of the pear cultivar 'Danxiahong'.Sci Data. 2025 Jun 18;12(1):1024. doi: 10.1038/s41597-025-05380-3. Sci Data. 2025. PMID: 40533474 Free PMC article.
-
CentIER: Accurate centromere identification for plant genomes.Plant Commun. 2024 Oct 14;5(10):101046. doi: 10.1016/j.xplc.2024.101046. Epub 2024 Aug 8. Plant Commun. 2024. PMID: 39118326 Free PMC article. No abstract available.
-
Pangenome analysis of Liriodendron reveals presence/absence variations associated with growth traits.BMC Plant Biol. 2025 Aug 8;25(1):1039. doi: 10.1186/s12870-025-07109-1. BMC Plant Biol. 2025. PMID: 40781705 Free PMC article.
References
-
- Ashwini D., Tiwari S. Use of CTAB method for isolation of good quality and quantity of DNA. J. Pure Appl. Microbiol. 2015;9:2271–2274.
-
- Audano P.A., Sulovari A., Graves-Lindsay T.A., Cantsilieris S., Sorensen M., Welch A.E., Dougherty M.L., Nelson B.J., Shah A., Dutcher S.K., et al. Characterizing the Major Structural Variant Alleles of the Human Genome. Cell. 2019;176:663–675.e19. doi: 10.1016/j.cell.2018.12.019. - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
