

Unconventional mechanism of action and resistance to rapalogs in renal cancer
- PMID: 38861592
- PMCID: PMC11194491
- DOI: 10.1073/pnas.2310793121
Unconventional mechanism of action and resistance to rapalogs in renal cancer
Abstract
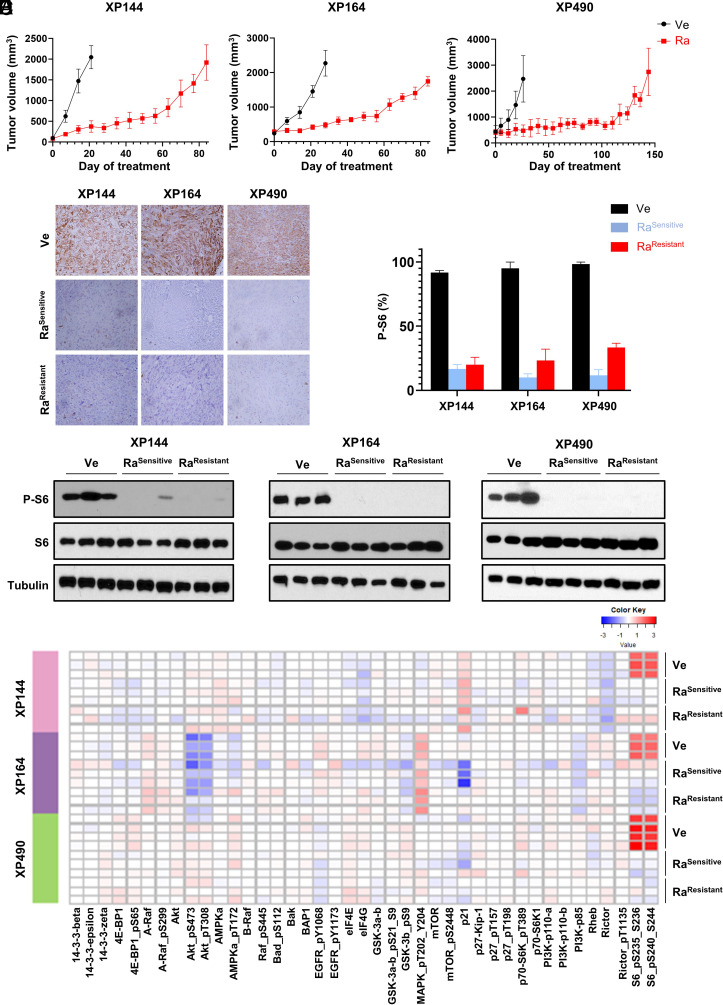
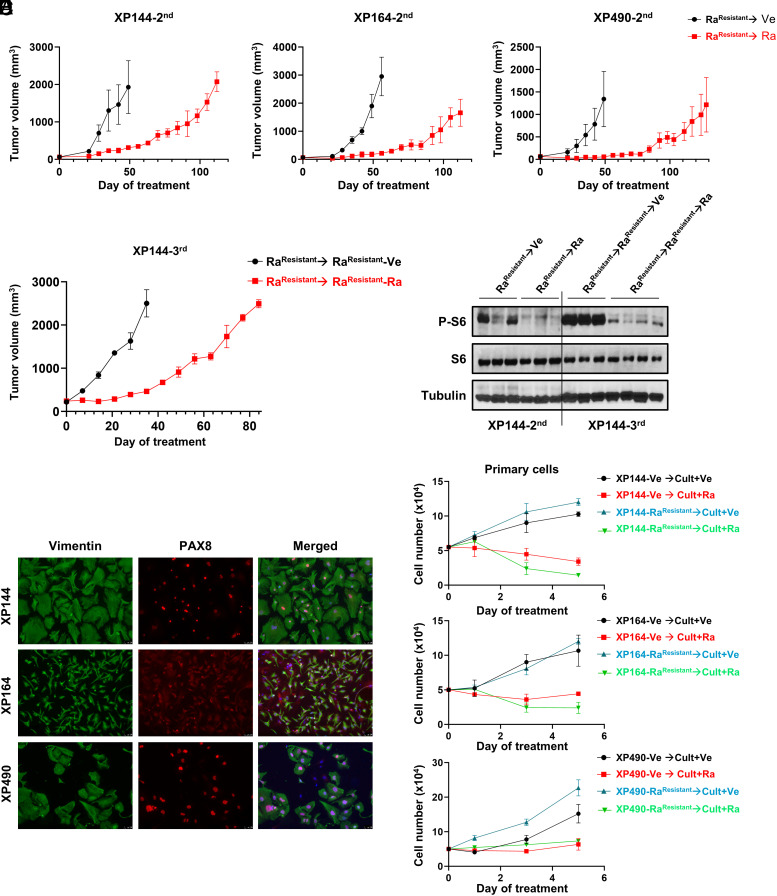
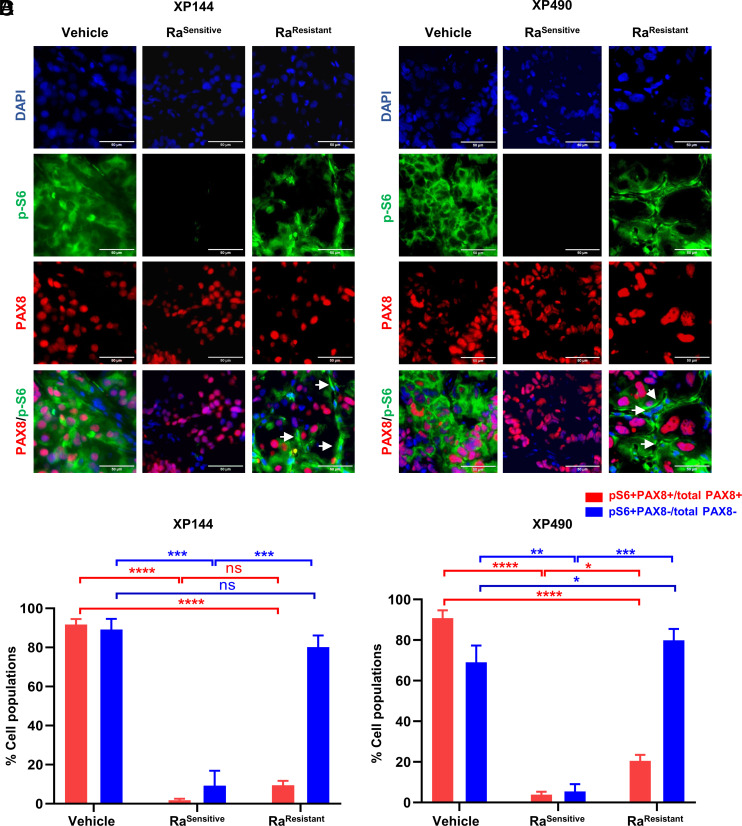
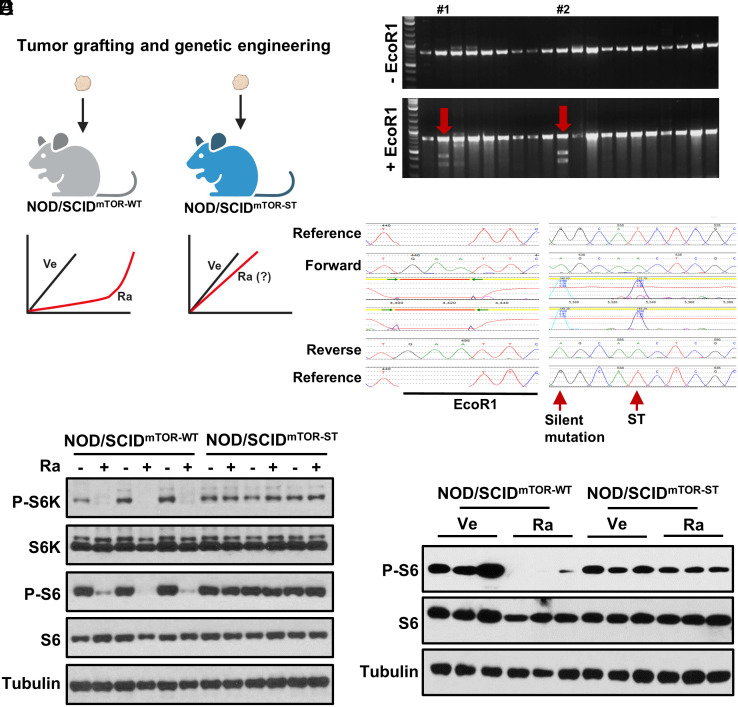
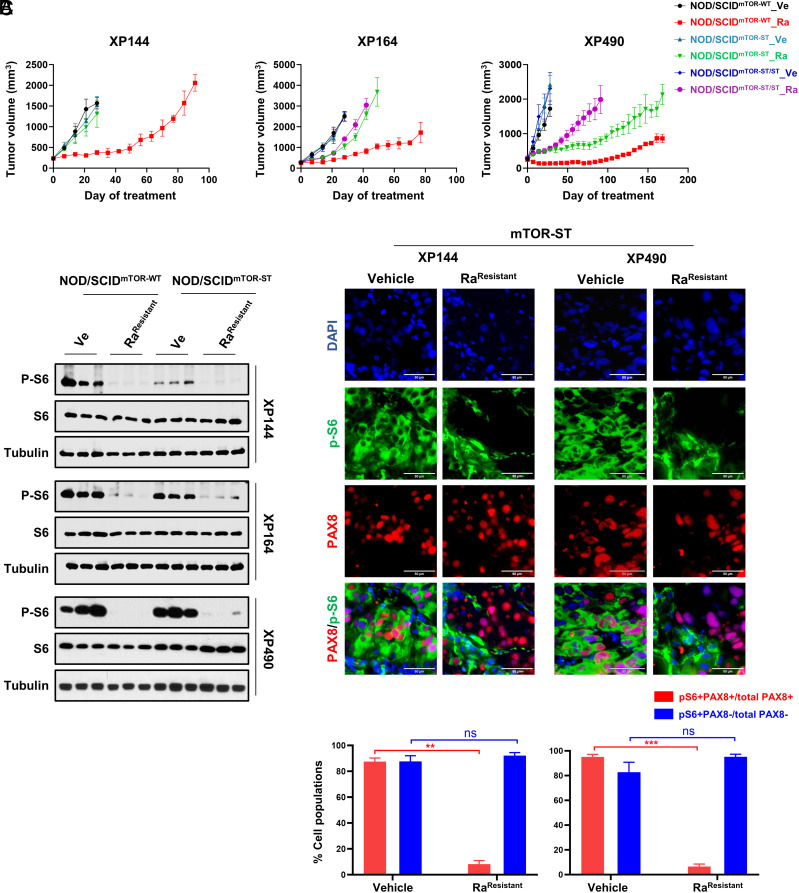
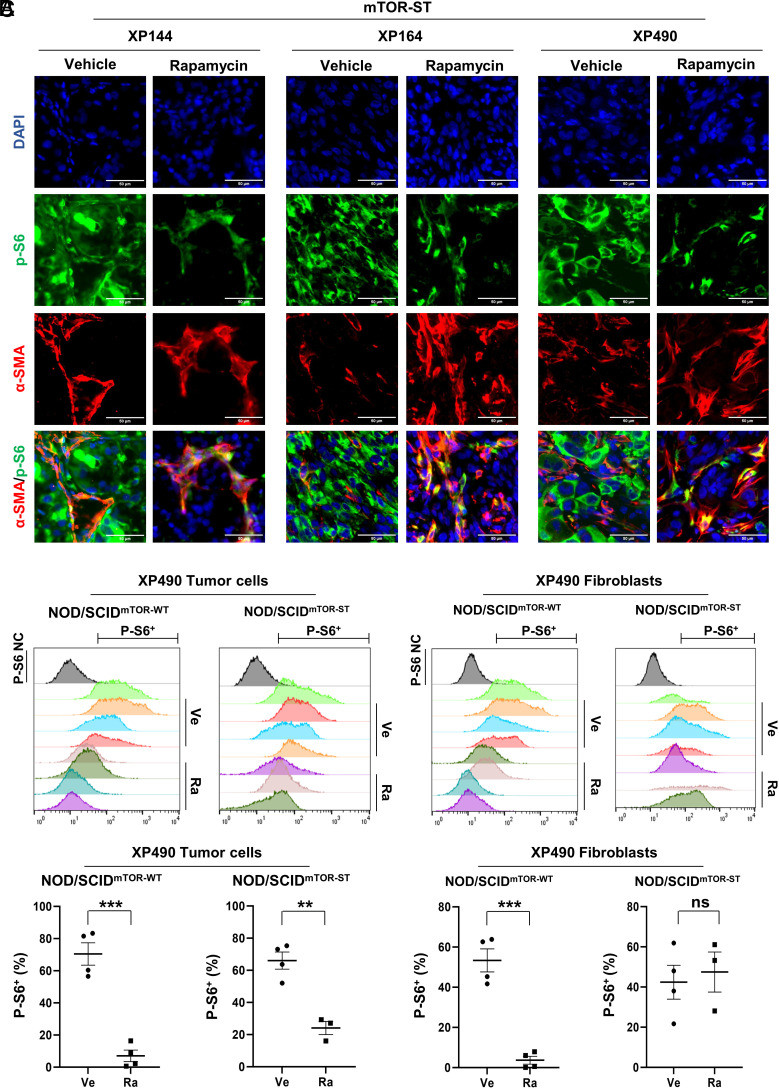
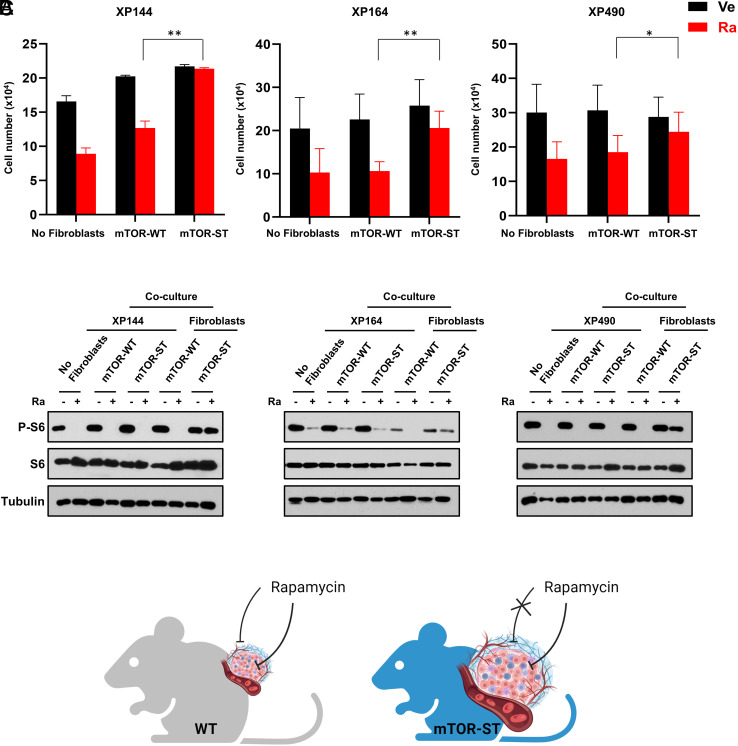
mTORC1 is aberrantly activated in renal cell carcinoma (RCC) and is targeted by rapalogs. As for other targeted therapies, rapalogs clinical utility is limited by the development of resistance. Resistance often results from target mutation, but mTOR mutations are rarely found in RCC. As in humans, prolonged rapalog treatment of RCC tumorgrafts (TGs) led to resistance. Unexpectedly, explants from resistant tumors became sensitive both in culture and in subsequent transplants in mice. Notably, resistance developed despite persistent mTORC1 inhibition in tumor cells. In contrast, mTORC1 became reactivated in the tumor microenvironment (TME). To test the role of the TME, we engineered immunocompromised recipient mice with a resistance mTOR mutation (S2035T). Interestingly, TGs became resistant to rapalogs in mTORS2035T mice. Resistance occurred despite mTORC1 inhibition in tumor cells and could be induced by coculturing tumor cells with mutant fibroblasts. Thus, enforced mTORC1 activation in the TME is sufficient to confer resistance to rapalogs. These studies highlight the importance of mTORC1 inhibition in nontumor cells for rapalog antitumor activity and provide an explanation for the lack of mTOR resistance mutations in RCC patients.
Keywords: Cancer-associated fibroblasts (CAFs); everolimus; kinase inhibitors; patient-derived xenograft (PDX); temsirolimus.
Conflict of interest statement
Competing interests statement:A patent application has been filed pertaining to aspects of this work.
Figures
References
-
- Lopez-Beltran A., Scarpelli M., Montironi R., Kirkali Z., 2004 WHO classification of the renal tumors of the adults. Eur. Urol. 49, 798–805 (2006). - PubMed
-
- Latif F., et al. , Identification of the von Hippel-Lindau disease tumor suppressor gene. Science 260, 1317–1320 (1993). - PubMed
-
- Maxwell P. H., et al. , The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 399, 271–275 (1999). - PubMed
-
- Ivan M., et al. , HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science 292, 464–468 (2001). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
