

Vibrio MARTX toxin processing and degradation of cellular Rab GTPases by the cytotoxic effector Makes Caterpillars Floppy
- PMID: 38861595
- PMCID: PMC11194500
- DOI: 10.1073/pnas.2316143121
Vibrio MARTX toxin processing and degradation of cellular Rab GTPases by the cytotoxic effector Makes Caterpillars Floppy
Abstract
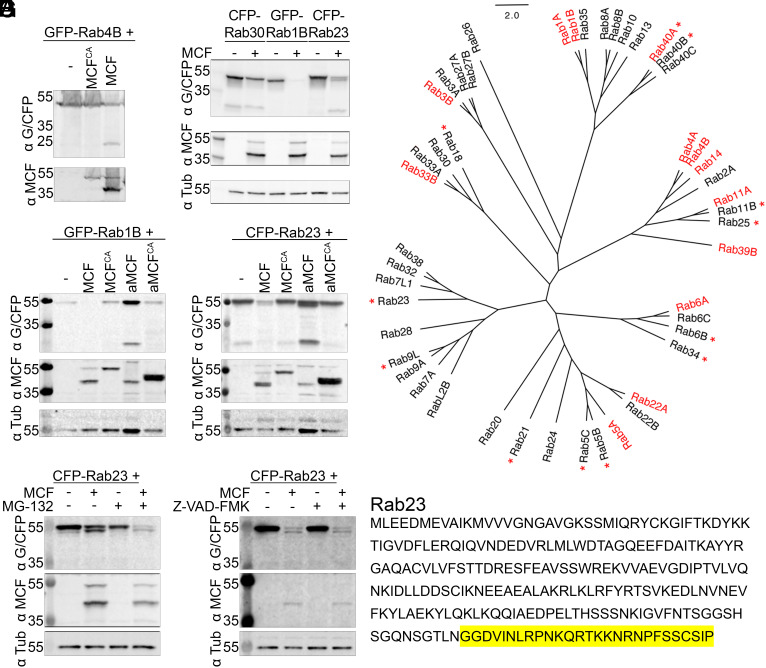
Vibrio vulnificus causes life-threatening wound and gastrointestinal infections, mediated primarily by the production of a Multifunctional-Autoprocessing Repeats-In-Toxin (MARTX) toxin. The most commonly present MARTX effector domain, the Makes Caterpillars Floppy-like (MCF) toxin, is a cysteine protease stimulated by host adenosine diphosphate (ADP) ribosylation factors (ARFs) to autoprocess. Here, we show processed MCF then binds and cleaves host
Keywords: MARTX toxin; Rab GTPases; Vibrio vulnificus; effector; host-pathogen interaction.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures






Update of
-
Vibrio MARTX toxin processing and degradation of cellular Rab GTPases by the cytotoxic effector Makes Caterpillars Floppy.bioRxiv [Preprint]. 2023 May 12:2023.04.19.537381. doi: 10.1101/2023.04.19.537381. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2316143121. doi: 10.1073/pnas.2316143121. PMID: 37131655 Free PMC article. Updated. Preprint.
Similar articles
-
Vibrio MARTX toxin processing and degradation of cellular Rab GTPases by the cytotoxic effector Makes Caterpillars Floppy.bioRxiv [Preprint]. 2023 May 12:2023.04.19.537381. doi: 10.1101/2023.04.19.537381. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2316143121. doi: 10.1073/pnas.2316143121. PMID: 37131655 Free PMC article. Updated. Preprint.
-
N-terminal autoprocessing and acetylation of multifunctional-autoprocessing repeats-in-toxins (MARTX) Makes Caterpillars Floppy-like effector is stimulated by adenosine diphosphate (ADP)-Ribosylation Factor 1 in advance of Golgi fragmentation.Cell Microbiol. 2020 Feb;22(2):e13133. doi: 10.1111/cmi.13133. Epub 2019 Nov 15. Cell Microbiol. 2020. PMID: 31658406 Free PMC article.
-
Cross-Kingdom Activation of Vibrio Toxins by ADP-Ribosylation Factor Family GTPases.J Bacteriol. 2020 Nov 19;202(24):e00278-20. doi: 10.1128/JB.00278-20. Print 2020 Nov 19. J Bacteriol. 2020. PMID: 32900828 Free PMC article. Review.
-
Makes caterpillars floppy-like effector-containing MARTX toxins require host ADP-ribosylation factor (ARF) proteins for systemic pathogenicity.Proc Natl Acad Sci U S A. 2019 Sep 3;116(36):18031-18040. doi: 10.1073/pnas.1905095116. Epub 2019 Aug 19. Proc Natl Acad Sci U S A. 2019. PMID: 31427506 Free PMC article.
-
Non-Repeat Segment 1 in Vibrio vulnificus MARTX toxin, which binds to biantennary N-glycans, is essential for host cell blebbing but dispensable for effector translocation.Microbiol Res. 2025 Jun;295:128108. doi: 10.1016/j.micres.2025.128108. Epub 2025 Feb 19. Microbiol Res. 2025. PMID: 39987752 Review.
Cited by
-
Vibrio MARTX toxin binding of biantennary N-glycans at host cell surfaces.Sci Adv. 2025 Apr 11;11(15):eadt0063. doi: 10.1126/sciadv.adt0063. Epub 2025 Apr 9. Sci Adv. 2025. PMID: 40203092 Free PMC article.
-
Biantennary N-glycans As Receptors for MARTX Toxins in Vibrio Pathogenesis.bioRxiv [Preprint]. 2024 Sep 12:2024.09.12.611726. doi: 10.1101/2024.09.12.611726. bioRxiv. 2024. PMID: 39314294 Free PMC article. Preprint.
-
Structure and activation mechanism of the Makes caterpillars floppy 1 toxin.Nat Commun. 2023 Dec 12;14(1):8226. doi: 10.1038/s41467-023-44069-2. Nat Commun. 2023. PMID: 38086871 Free PMC article.
References
-
- Satchell K. J. F., Multifunctional-autoprocessing repeats-in-toxin (MARTX) Toxins of Vibrios. Microbiol. Spectr. 3, (2015), 10.1128/microbiolspec.ve-0002-2014. - DOI - PMC - PubMed
-
- Herrera A., et al. , N-terminal autoprocessing and acetylation of multifunctional-autoprocessing repeats-in-toxins (MARTX) Makes Caterpillars Floppy-like effector is stimulated by adenosine diphosphate (ADP)-Ribosylation Factor 1 in advance of Golgi fragmentation. Cell Microbiol. 22, e13133 (2020). - PMC - PubMed
MeSH terms
Substances
Grants and funding
- CCSG P30 CA060553/HHS | NIH | National Cancer Institute (NCI)
- S10OD025194/HHS | NIH | NIH Office of the Director (OD)
- P41 GM108569/GM/NIGMS NIH HHS/United States
- R01 GM129325/GM/NIGMS NIH HHS/United States
- F31 AI172382/AI/NIAID NIH HHS/United States
- HHSN272201700060C/AI/NIAID NIH HHS/United States
- 75N93022C00035/AI/NIAID NIH HHS/United States
- R01-GM129325/HHS | National Institutes of Health (NIH)
- 085P1000817/Michigan Economic Development Corporation (MEDC)
- K99 GM143571/HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- DE-AC02- 06CH11357/U.S. Department of Energy (DOE)
- P30 CA060553/CA/NCI NIH HHS/United States
- R37 AI092825/AI/NIAID NIH HHS/United States
- S10 OD025194/OD/NIH HHS/United States
- K99 AI167819/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
