

Functional analysis of the Aspergillus fumigatus kinome identifies a druggable DYRK kinase that regulates septal plugging
- PMID: 38862481
- PMCID: PMC11166925
- DOI: 10.1038/s41467-024-48592-8
Functional analysis of the Aspergillus fumigatus kinome identifies a druggable DYRK kinase that regulates septal plugging
Erratum in
-
Author Correction: Functional analysis of the Aspergillus fumigatus kinome identifies a druggable DYRK kinase that regulates septal plugging.Nat Commun. 2024 Aug 20;15(1):7126. doi: 10.1038/s41467-024-50129-y. Nat Commun. 2024. PMID: 39164238 Free PMC article. No abstract available.
Abstract
More than 10 million people suffer from lung diseases caused by the pathogenic fungus Aspergillus fumigatus. Azole antifungals represent first-line therapeutics for most of these infections but resistance is rising, therefore the identification of antifungal targets whose inhibition synergises with the azoles could improve therapeutic outcomes. Here, we generate a library of 111 genetically barcoded null mutants of Aspergillus fumigatus in genes encoding protein kinases, and show that loss of function of kinase YakA results in hypersensitivity to the azoles and reduced pathogenicity. YakA is an orthologue of Candida albicans Yak1, a TOR signalling pathway kinase involved in modulation of stress responsive transcriptional regulators. We show that YakA has been repurposed in A. fumigatus to regulate blocking of the septal pore upon exposure to stress. Loss of YakA function reduces the ability of A. fumigatus to penetrate solid media and to grow in mouse lung tissue. We also show that 1-ethoxycarbonyl-beta-carboline (1-ECBC), a compound previously shown to inhibit C. albicans Yak1, prevents stress-mediated septal spore blocking and synergises with the azoles to inhibit A. fumigatus growth.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
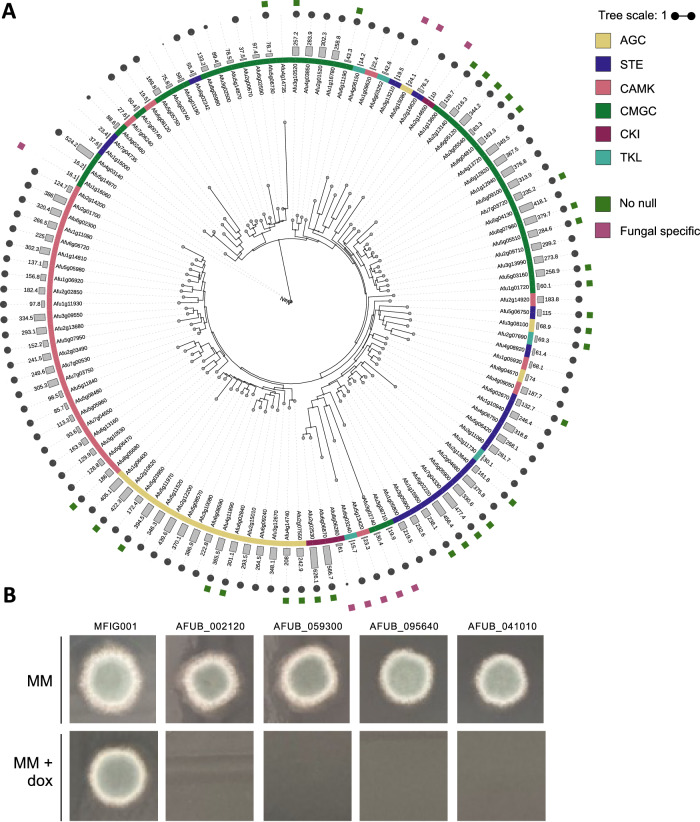

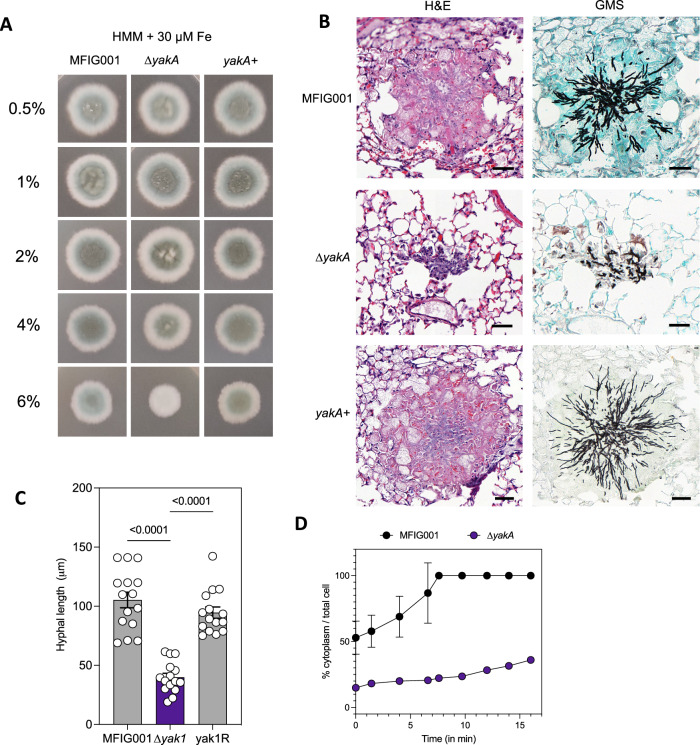



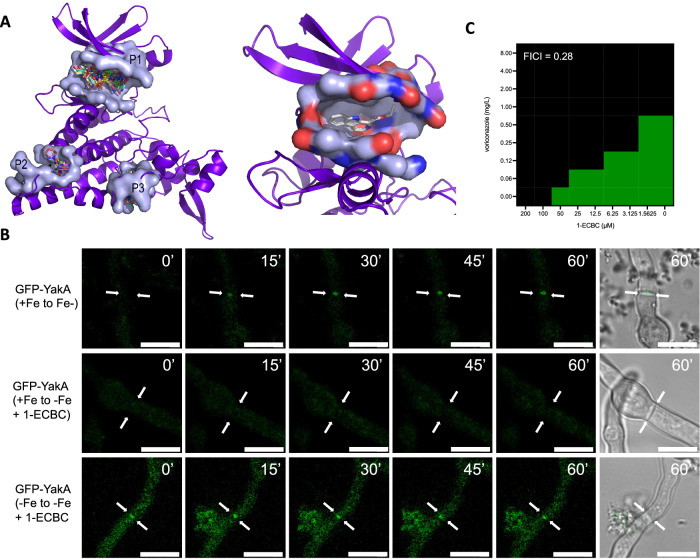
Update of
-
Functional analysis of the Aspergillus fumigatus kinome reveals a DYRK kinase involved in septal plugging is a novel antifungal drug target.Res Sq [Preprint]. 2023 May 30:rs.3.rs-2960526. doi: 10.21203/rs.3.rs-2960526/v1. Res Sq. 2023. Update in: Nat Commun. 2024 Jun 11;15(1):4984. doi: 10.1038/s41467-024-48592-8. PMID: 37398159 Free PMC article. Updated. Preprint.
References
-
- Brown, G. D. et al. Hidden killers: human fungal infections. Sci. Transl. Med.4, 165rv13 (2012). - PubMed
-
- Alastruey-Izquierdo, A. & World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action (World Health Organization, 2022).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
