

Kinetic and dynamical properties of truncated hemoglobins of the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125
- PMID: 38864722
- PMCID: PMC11168075
- DOI: 10.1002/pro.5064
Kinetic and dynamical properties of truncated hemoglobins of the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125
Abstract
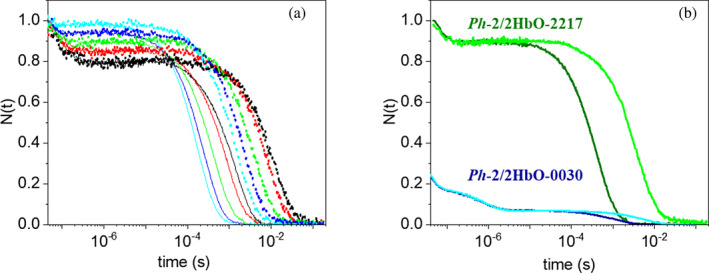
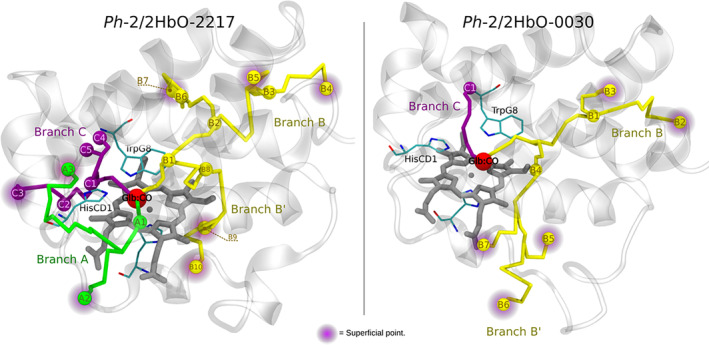
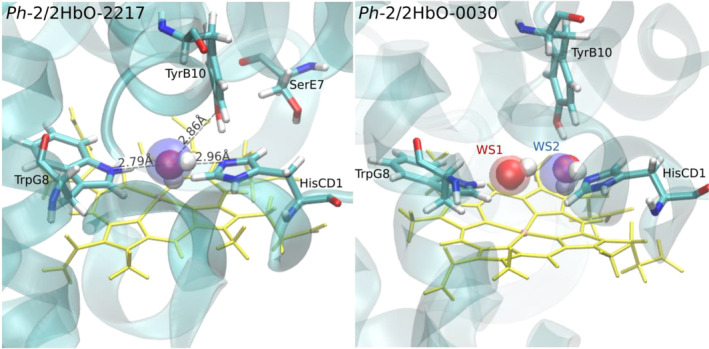
Due to the low temperature, the Antarctic marine environment is challenging for protein functioning. Cold-adapted organisms have evolved proteins endowed with higher flexibility and lower stability in comparison to their thermophilic homologs, resulting in enhanced reaction rates at low temperatures. The Antarctic bacterium Pseudoalteromonas haloplanktis TAC125 (PhTAC125) genome is one of the few examples of coexistence of multiple hemoglobin genes encoding, among others, two constitutively transcribed 2/2 hemoglobins (2/2Hbs), also named truncated Hbs (TrHbs), belonging to the Group II (or O), annotated as PSHAa0030 and PSHAa2217. In this work, we describe the ligand binding kinetics and their interrelationship with the dynamical properties of globin Ph-2/2HbO-2217 by combining experimental and computational approaches and implementing a new computational method to retrieve information from molecular dynamic trajectories. We show that our approach allows us to identify docking sites within the protein matrix that are potentially able to transiently accommodate ligands and migration pathways connecting them. Consistently with ligand rebinding studies, our modeling suggests that the distal heme pocket is connected to the solvent through a low energy barrier, while inner cavities play only a minor role in modulating rebinding kinetics.
Keywords: CO rebinding kinetics; bacterial globin; cold adaptation; molecular dynamic simulations; oxidative/nitrosative stress.
© 2024 The Protein Society.
Figures








Similar articles
-
The globins of cold-adapted Pseudoalteromonas haloplanktis TAC125: from the structure to the physiological functions.Adv Microb Physiol. 2013;63:329-89. doi: 10.1016/B978-0-12-407693-8.00008-X. Adv Microb Physiol. 2013. PMID: 24054800 Review.
-
Coexistence of multiple globin genes conferring protection against nitrosative stress to the Antarctic bacterium Pseudoalteromonas haloplanktis TAC125.Nitric Oxide. 2018 Feb 28;73:39-51. doi: 10.1016/j.niox.2017.12.006. Epub 2017 Dec 21. Nitric Oxide. 2018. PMID: 29275194
-
Cold-Adaptation Signatures in the Ligand Rebinding Kinetics to the Truncated Hemoglobin of the Antarctic Bacterium Pseudoalteromonas haloplanktis TAC125.J Phys Chem B. 2018 Dec 13;122(49):11649-11661. doi: 10.1021/acs.jpcb.8b07682. Epub 2018 Oct 3. J Phys Chem B. 2018. PMID: 30230844
-
Structural flexibility of the heme cavity in the cold-adapted truncated hemoglobin from the Antarctic marine bacterium Pseudoalteromonas haloplanktis TAC125.FEBS J. 2015 Aug;282(15):2948-65. doi: 10.1111/febs.13335. Epub 2015 Jun 26. FEBS J. 2015. PMID: 26040838
-
Conformational Flexibility Drives Cold Adaptation in Pseudoalteromonas haloplanktis TAC125 Globins.Antioxid Redox Signal. 2020 Feb 20;32(6):396-411. doi: 10.1089/ars.2019.7887. Epub 2019 Nov 11. Antioxid Redox Signal. 2020. PMID: 31578873 Review.
References
-
- Abbruzzetti S, Grandi E, Bruno S, Faggiano S, Spyrakis F, Mozzarelli A, et al. Ligand migration in nonsymbiotic hemoglobin AHb1 from Arabidopsis thaliana . J Phys Chem B. 2007;111:12582–12590. - PubMed
-
- Abbruzzetti S, He C, Ogata H, Bruno S, Viappiani C, Knipp M. Heterogeneous kinetics of the carbon monoxide association and dissociation reaction to nitrophorin 4 and 7 coincide with structural heterogeneity of the gate‐loop. J Am Chem Soc. 2012;134:9986–9998. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
