

Shoot hydraulic impairments induced by root waterlogging: Parallels and contrasts with drought
- PMID: 38865443
- PMCID: PMC11663564
- DOI: 10.1093/plphys/kiae336
Shoot hydraulic impairments induced by root waterlogging: Parallels and contrasts with drought
Abstract
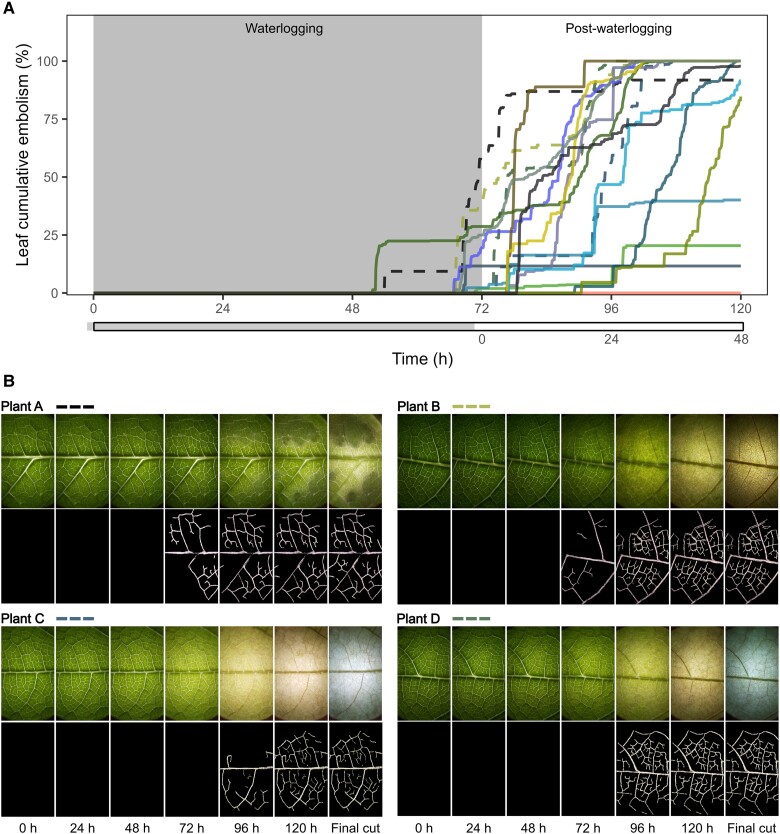
Soil waterlogging and drought correspond to contrasting water extremes resulting in plant dehydration. Dehydration in response to waterlogging occurs due to impairments to root water transport, but no previous study has addressed whether limitations to water transport occur beyond this organ or whether dehydration alone can explain shoot impairments. Using common bean (Phaseolus vulgaris) as a model species, we report that waterlogging also impairs water transport in leaves and stems. During the very first hours of waterlogging, leaves transiently dehydrated to water potentials close to the turgor loss point, possibly driving rapid stomatal closure and partially explaining the decline in leaf hydraulic conductance. The initial decline in leaf hydraulic conductance (occurring within 24 h), however, surpassed the levels predicted to occur based solely on dehydration. Constraints to leaf water transport resulted in a hydraulic disconnection between leaves and stems, furthering leaf dehydration during waterlogging and after soil drainage. As leaves dehydrated later during waterlogging, leaf embolism initiated and extensive embolism levels amplified leaf damage. The hydraulic disconnection between leaves and stems prevented stem water potentials from declining below the threshold for critical embolism levels in response to waterlogging. This allowed plants to survive waterlogging and soil drainage. In summary, leaf and stem dehydration are central in defining plant impairments in response to waterlogging, thus creating similarities between waterlogging and drought. Yet, our findings point to the existence of additional players (likely chemicals) partially controlling the early declines in leaf hydraulic conductance and contributing to leaf damage during waterlogging.
© The Author(s) 2024. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures




Similar articles
-
Dynamic soil hydraulic resistance regulates stomata.New Phytol. 2024 Oct;244(1):147-158. doi: 10.1111/nph.20020. Epub 2024 Aug 2. New Phytol. 2024. PMID: 39096020
-
Neither xylem collapse, cavitation, or changing leaf conductance drive stomatal closure in wheat.Plant Cell Environ. 2020 Apr;43(4):854-865. doi: 10.1111/pce.13722. Epub 2020 Feb 9. Plant Cell Environ. 2020. PMID: 31953855
-
Coordination between leaf, stem, and root hydraulics and gas exchange in three arid-zone angiosperms during severe drought and recovery.Plant Cell Environ. 2018 Dec;41(12):2869-2881. doi: 10.1111/pce.13418. Epub 2018 Sep 14. Plant Cell Environ. 2018. PMID: 30106477
-
Water relations in tree physiology: where to from here?Tree Physiol. 2017 Jan 31;37(1):18-32. doi: 10.1093/treephys/tpw102. Tree Physiol. 2017. PMID: 28173481 Review.
-
Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept.Plant Cell Environ. 2017 Jun;40(6):962-976. doi: 10.1111/pce.12846. Epub 2016 Dec 14. Plant Cell Environ. 2017. PMID: 27739594 Review.
Cited by
-
Between Two Extremes: Tripsacum dactyloides Root Anatomical Responses to Drought and Waterlogging.Plant Direct. 2025 Jul 19;9(7):e70097. doi: 10.1002/pld3.70097. eCollection 2025 Jul. Plant Direct. 2025. PMID: 40687876 Free PMC article.
-
Hypoxia as challenge and opportunity: From cells to crops, to synthetic biology.Plant Physiol. 2024 Dec 24;197(1):kiae640. doi: 10.1093/plphys/kiae640. Plant Physiol. 2024. PMID: 39657271 Free PMC article. No abstract available.
References
-
- Bourbia I, Brodribb TJ. A new technique for monitoring plant transpiration under field conditions using leaf optical dendrometry. Agric For Meteorol. 2023:331:109328. 10.1016/j.agrformet.2023.109328 - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
