

Genome-wide profiles of H3K9me3, H3K27me3 modifications, and DNA methylation during diapause of Asian corn borer (Ostrinia furnacalis)
- PMID: 38866549
- PMCID: PMC11216315
- DOI: 10.1101/gr.278661.123
Genome-wide profiles of H3K9me3, H3K27me3 modifications, and DNA methylation during diapause of Asian corn borer (Ostrinia furnacalis)
Abstract
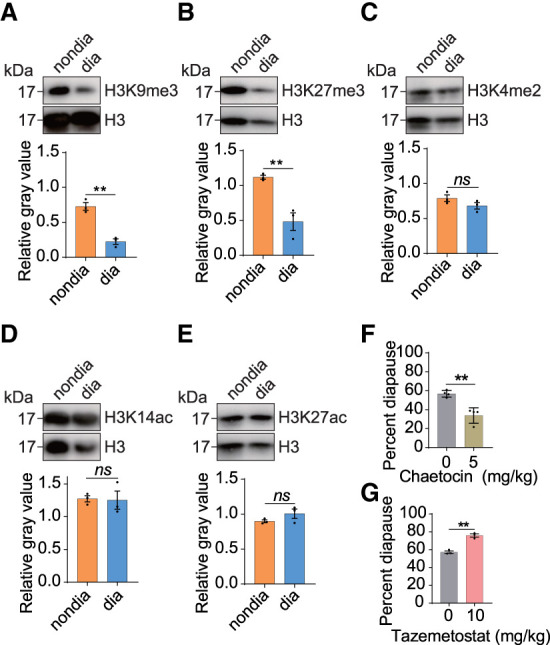
Diapause represents a crucial adaptive strategy used by insects to cope with changing environmental conditions. In North China, the Asian corn borer (Ostrinia furnacalis) enters a winter larval diapause stage. Although there is growing evidence implicating epigenetic mechanisms in diapause regulation, it remains unclear whether dynamic genome-wide profiles of epigenetic modifications exist during this process. By investigating multiple histone modifications, we have discovered the essential roles of H3K9me3 and H3K27me3 during diapause of the Asian corn borer. Building upon previous findings in vertebrates highlighting the connection between DNA methylation and repressive histone methylations, we have examined changes in the genome-wide profile of H3K9me3, H3K27me3, and DNA methylation at the nondiapause, prediapause, and diapause stages. Data analysis reveals significant alterations in these three modifications during diapause. Moreover, we observe a correlation between the H3K9me3 and H3K27me3 modification sites during diapause, whereas DNA modifications show little association with either H3K9me3 or H3K27me3. Integrative analysis of epigenome and expression data unveils the relationship between these epigenetic modifications and gene expression levels at corresponding diapause stages. Furthermore, by studying the function of histone modifications on genes known to be important in diapause, especially those involved in the juvenile pathway, we discover that the juvenile hormone pathway lies downstream from H3K9me3 and H3K27me3 histone modifications. Finally, the analysis of gene loci with modified modifications unreported in diapause uncovers novel pathways potentially crucial in diapause regulation. This study provides a valuable resource for future investigations aiming to elucidate the underlying mechanisms of diapause.
© 2024 Lv et al.; Published by Cold Spring Harbor Laboratory Press.
Figures







Similar articles
-
Genome-wide analysis reveals transcriptional and translational changes during diapause of the Asian corn borer (Ostrinia furnacalis).BMC Biol. 2024 Sep 13;22(1):206. doi: 10.1186/s12915-024-02000-1. BMC Biol. 2024. PMID: 39272107 Free PMC article.
-
Inheritance of photoperiodic control of larval diapause in the Asian corn borer Ostrinia furnacalis (Guenée).Bull Entomol Res. 2015 Jun;105(3):326-34. doi: 10.1017/S0007485315000140. Epub 2015 Mar 17. Bull Entomol Res. 2015. PMID: 25779483
-
EXPRESSION PATTERNS OF THE GLYCOGEN PHOSPHORYLASE GENE RELATED TO LARVAL DIAPAUSE IN Ostrinia furnacalis.Arch Insect Biochem Physiol. 2016 Apr;91(4):210-20. doi: 10.1002/arch.21318. Epub 2016 Jan 8. Arch Insect Biochem Physiol. 2016. PMID: 26748939
-
Epigenomics in stress tolerance of plants under the climate change.Mol Biol Rep. 2023 Jul;50(7):6201-6216. doi: 10.1007/s11033-023-08539-6. Epub 2023 Jun 9. Mol Biol Rep. 2023. PMID: 37294468 Review.
-
Transgenerational inheritance: how impacts to the epigenetic and genetic information of parents affect offspring health.Hum Reprod Update. 2019 Sep 11;25(5):518-540. doi: 10.1093/humupd/dmz017. Hum Reprod Update. 2019. PMID: 31374565
Cited by
-
The PBAP chromatin remodeling complex mediates summer diapause via H3K4me3-driven juvenile hormone regulation in Colaphellus bowringi.Proc Natl Acad Sci U S A. 2025 Mar 25;122(12):e2422328122. doi: 10.1073/pnas.2422328122. Epub 2025 Mar 20. Proc Natl Acad Sci U S A. 2025. PMID: 40112108
-
Noncanonical action of circadian clock genes controls winter diapause entry via the NuA4/TIP60 complex in Harmonia axyridis.Proc Natl Acad Sci U S A. 2025 Jul 15;122(28):e2510550122. doi: 10.1073/pnas.2510550122. Epub 2025 Jul 10. Proc Natl Acad Sci U S A. 2025. PMID: 40638091
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources