

Three-dimensional chromatin reorganization regulates B cell development during ageing
- PMID: 38866970
- PMCID: PMC11178499
- DOI: 10.1038/s41556-024-01424-9
Three-dimensional chromatin reorganization regulates B cell development during ageing
Abstract
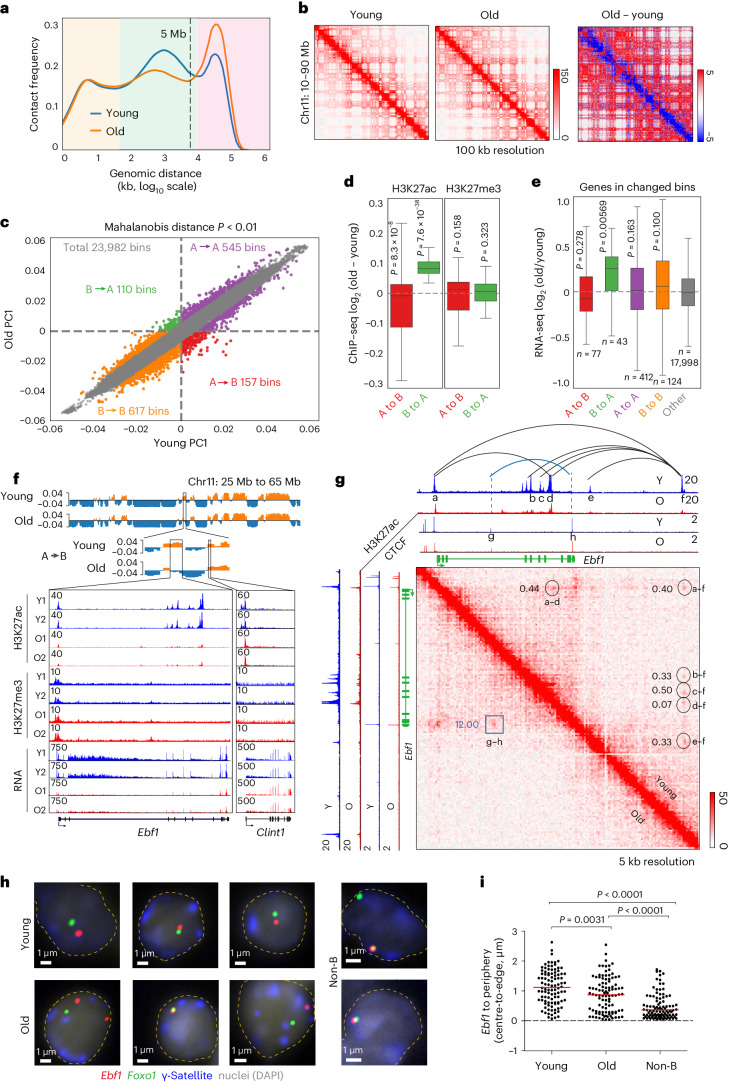
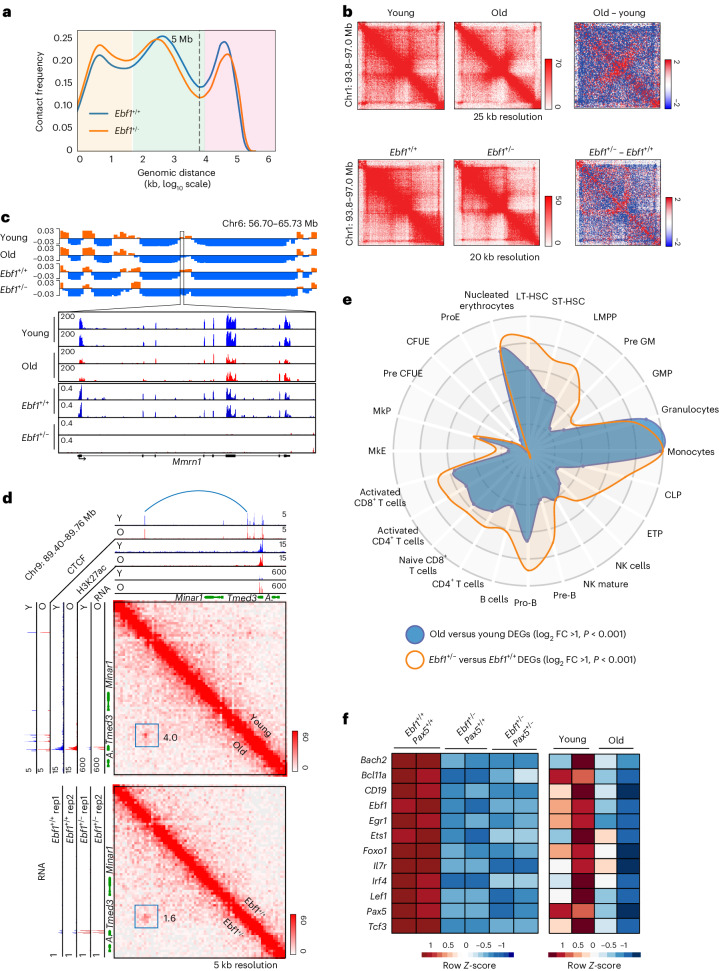
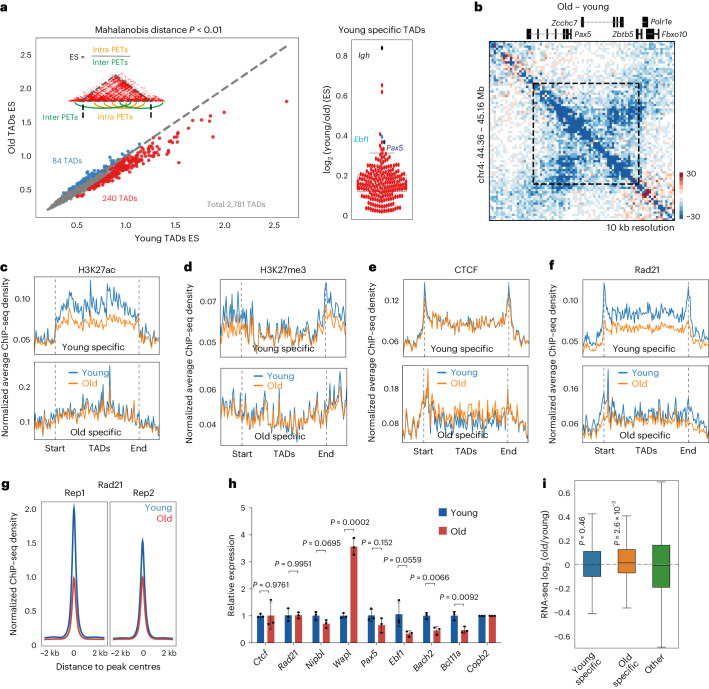
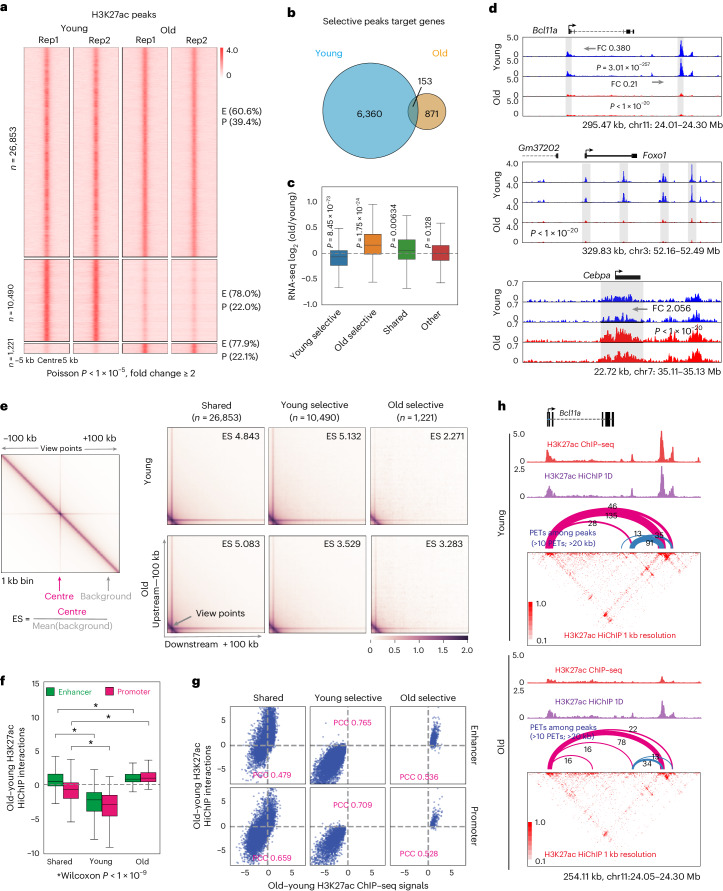
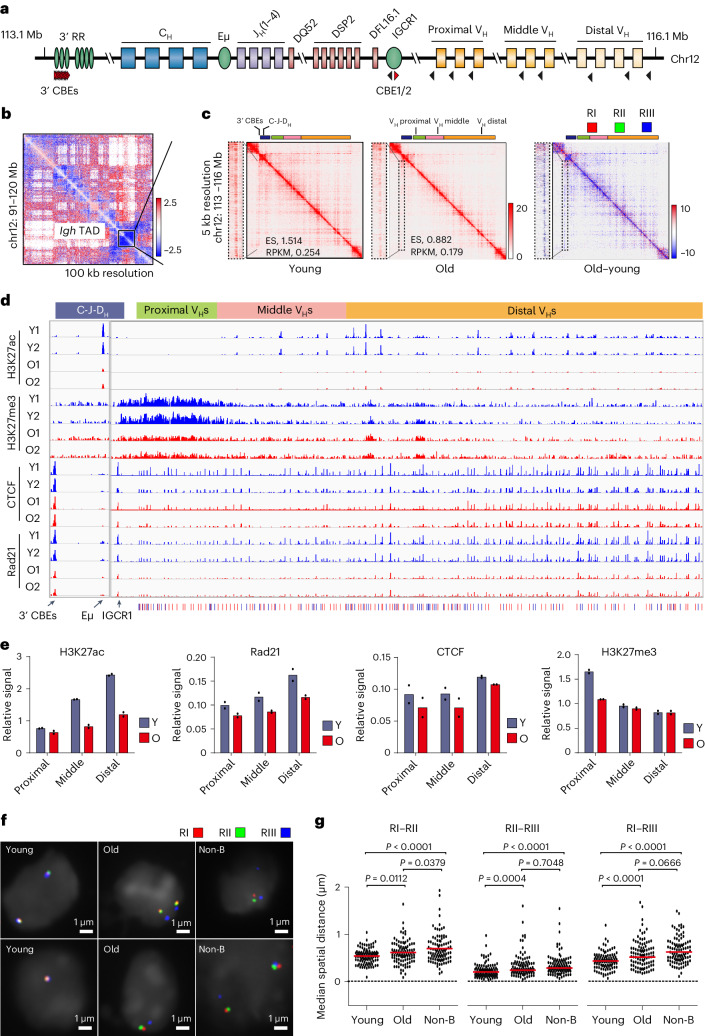
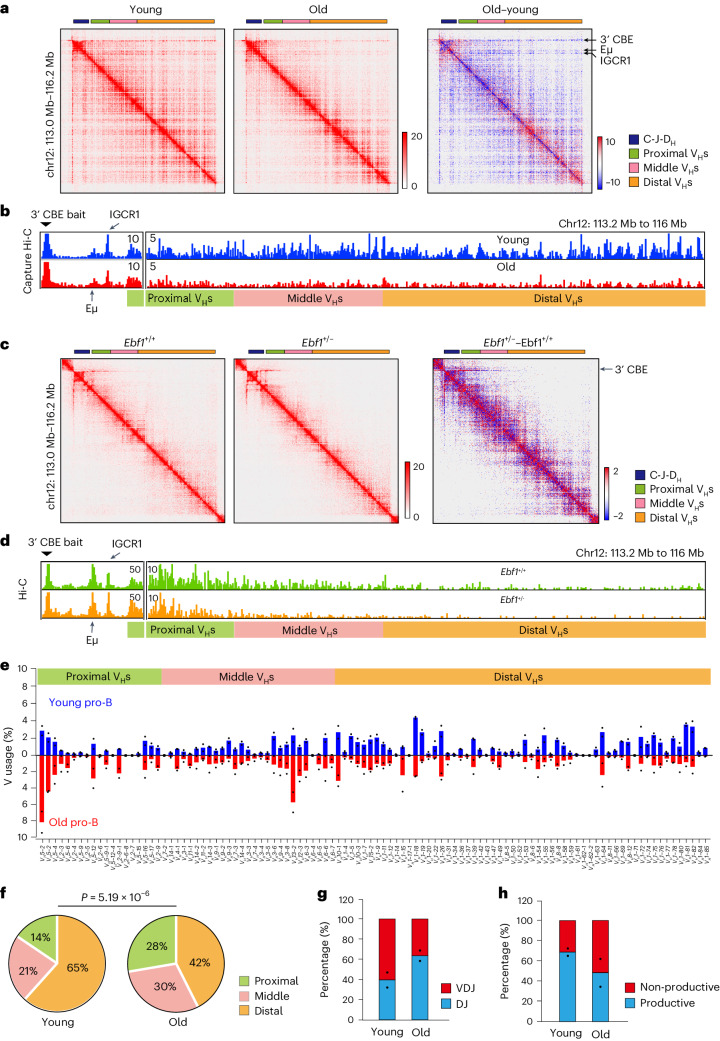
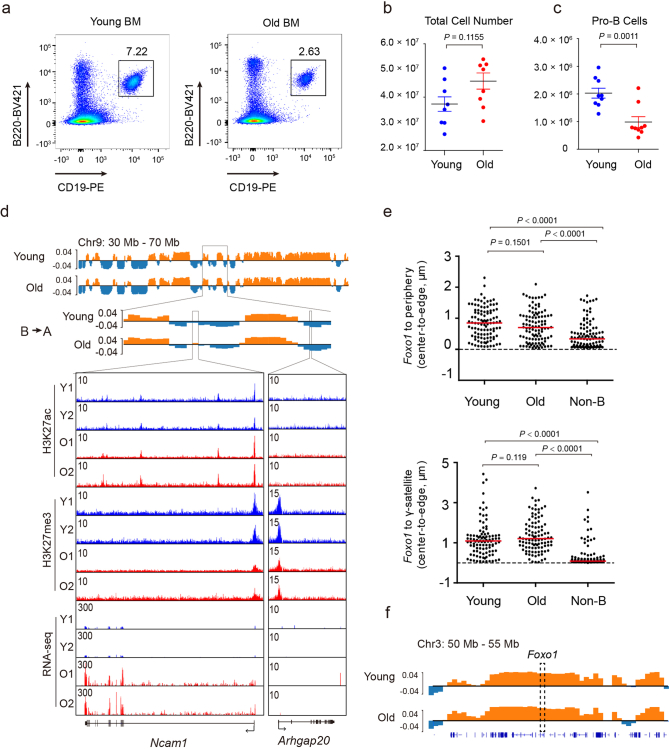
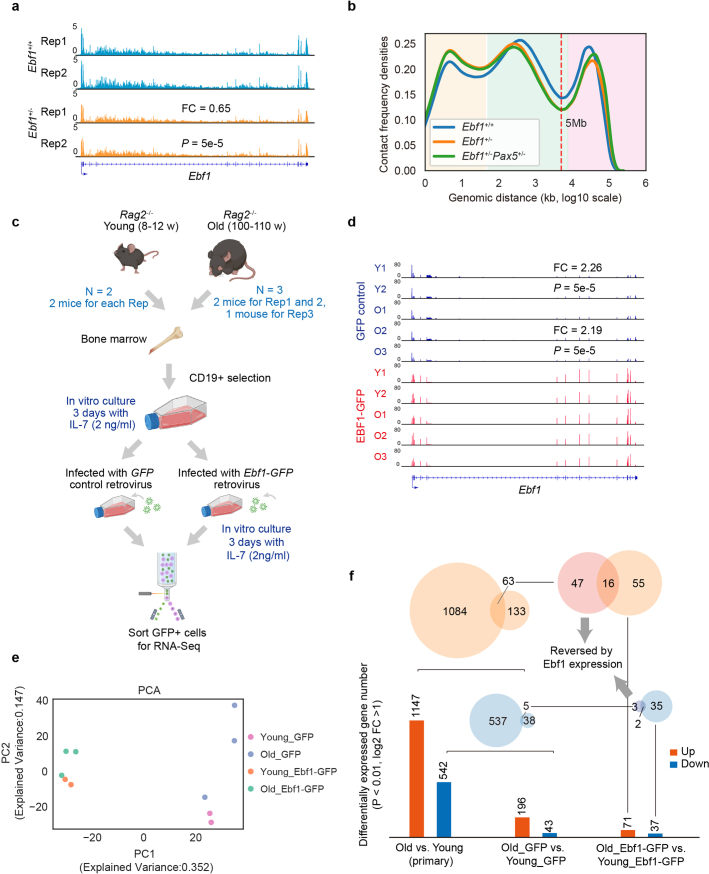
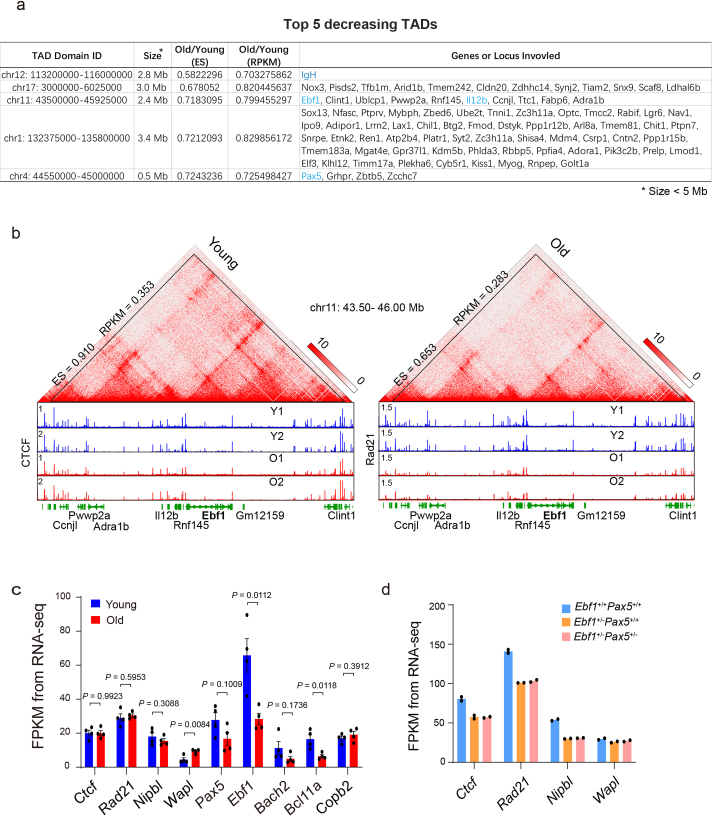
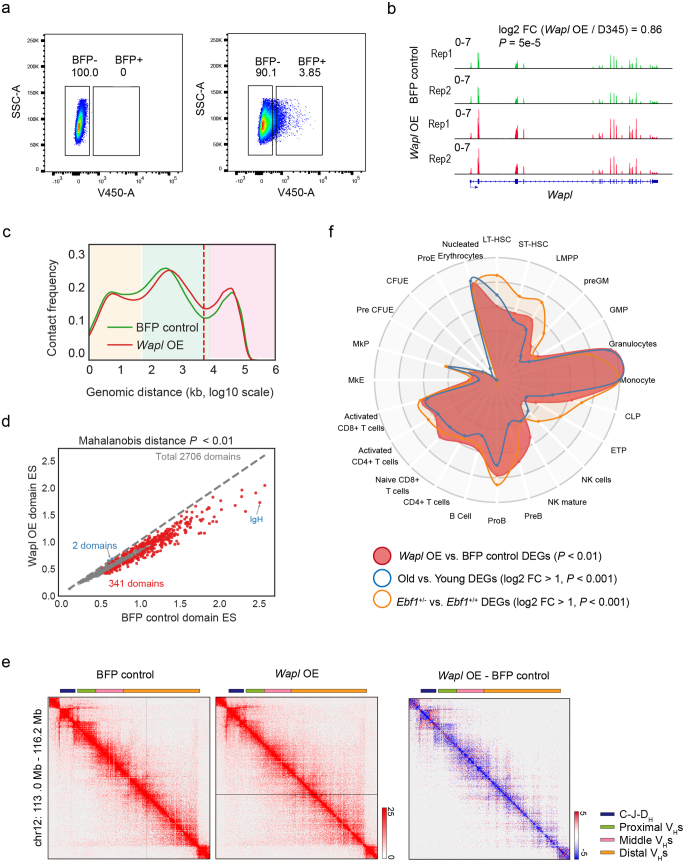
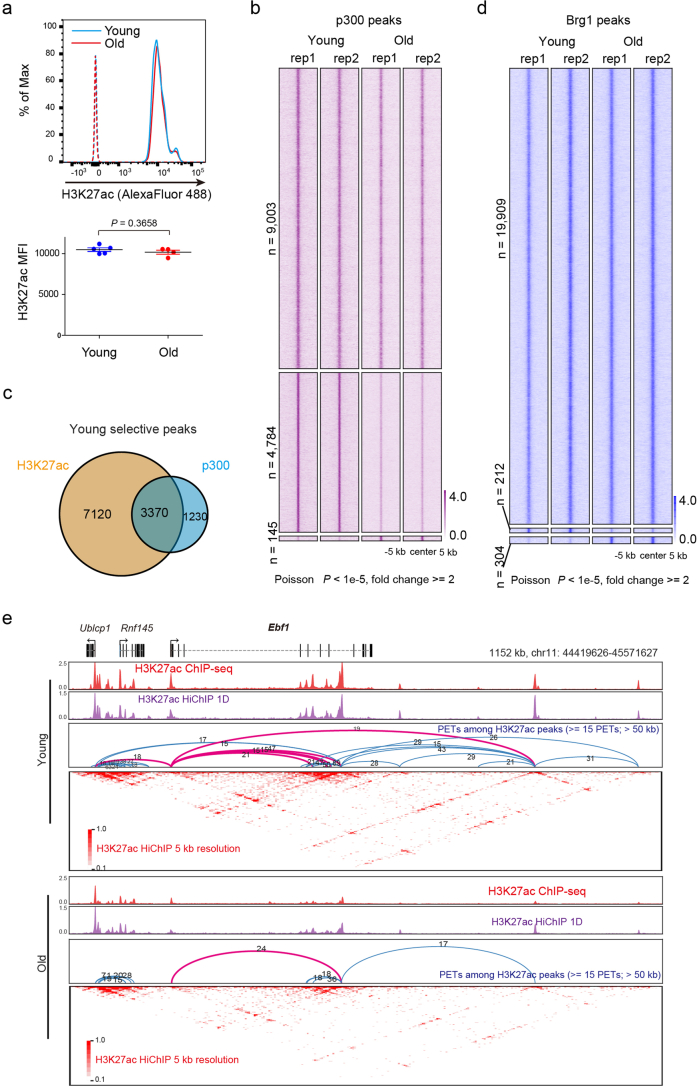
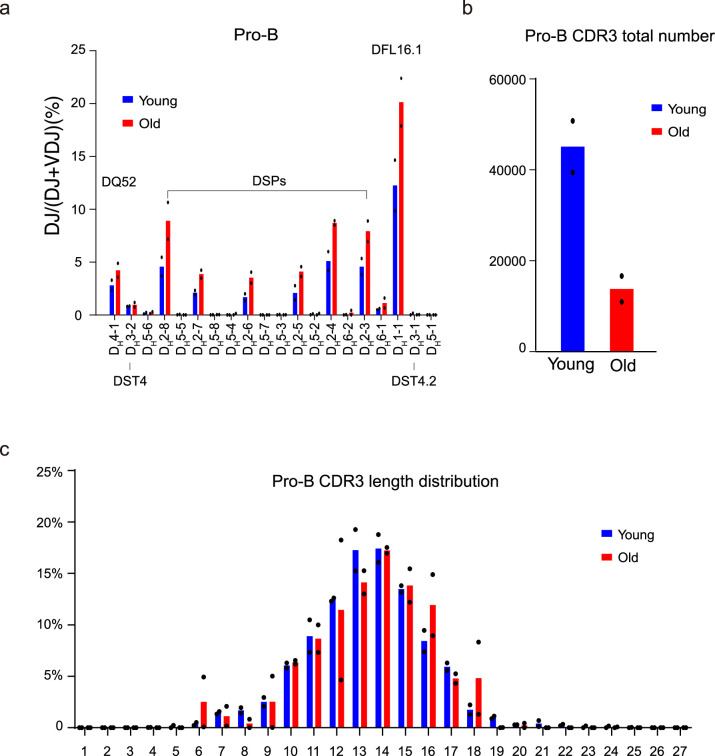
The contribution of three-dimensional genome organization to physiological ageing is not well known. Here we show that large-scale chromatin reorganization distinguishes young and old bone marrow progenitor (pro-) B cells. These changes result in increased interactions at the compartment level and reduced interactions within topologically associated domains (TADs). The gene encoding Ebf1, a key B cell regulator, switches from compartment A to B with age. Genetically reducing Ebf1 recapitulates some features of old pro-B cells. TADs that are most reduced with age contain genes important for B cell development, including the immunoglobulin heavy chain (Igh) locus. Weaker intra-TAD interactions at Igh correlate with altered variable (V), diversity (D) and joining (J) gene recombination. Our observations implicate three-dimensional chromatin reorganization as a major driver of pro-B cell phenotypes that impair B lymphopoiesis with age.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
