

DNA mismatch and damage patterns revealed by single-molecule sequencing
- PMID: 38867045
- PMCID: PMC11216816
- DOI: 10.1038/s41586-024-07532-8
DNA mismatch and damage patterns revealed by single-molecule sequencing
Abstract
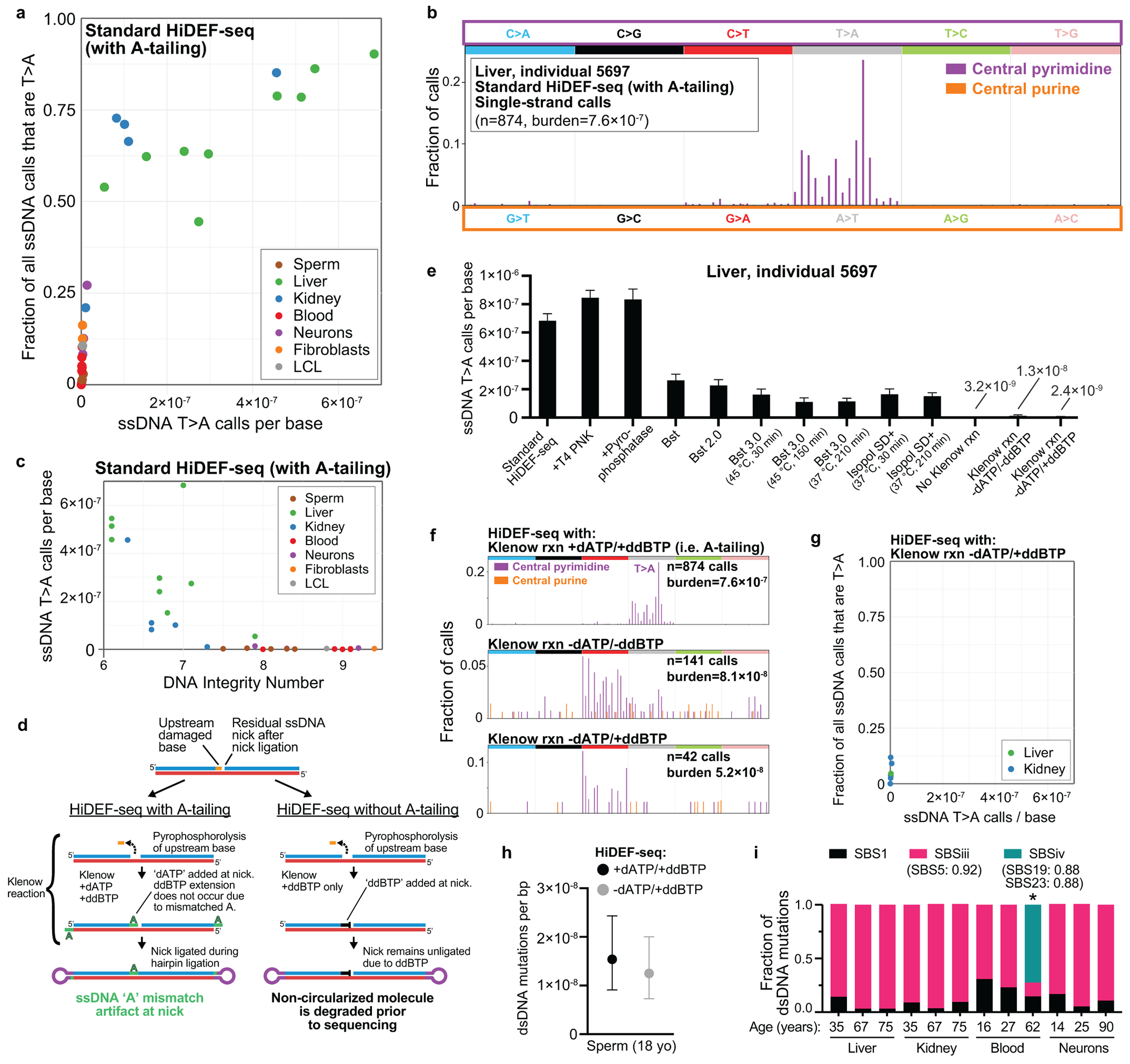
Mutations accumulate in the genome of every cell of the body throughout life, causing cancer and other diseases1,2. Most mutations begin as nucleotide mismatches or damage in one of the two strands of the DNA before becoming double-strand mutations if unrepaired or misrepaired3,4. However, current DNA-sequencing technologies cannot accurately resolve these initial single-strand events. Here we develop a single-molecule, long-read sequencing method (Hairpin Duplex Enhanced Fidelity sequencing (HiDEF-seq)) that achieves single-molecule fidelity for base substitutions when present in either one or both DNA strands. HiDEF-seq also detects cytosine deamination-a common type of DNA damage-with single-molecule fidelity. We profiled 134 samples from diverse tissues, including from individuals with cancer predisposition syndromes, and derive from them single-strand mismatch and damage signatures. We find correspondences between these single-strand signatures and known double-strand mutational signatures, which resolves the identity of the initiating lesions. Tumours deficient in both mismatch repair and replicative polymerase proofreading show distinct single-strand mismatch patterns compared to samples that are deficient in only polymerase proofreading. We also define a single-strand damage signature for APOBEC3A. In the mitochondrial genome, our findings support a mutagenic mechanism occurring primarily during replication. As double-strand DNA mutations are only the end point of the mutation process, our approach to detect the initiating single-strand events at single-molecule resolution will enable studies of how mutations arise in a variety of contexts, especially in cancer and ageing.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing Interests
A patent application for HiDEF-seq has been filed (G.D.E.). G.D.E. owns equity in DNA sequencing companies (Illumina, Oxford Nanopore Technologies, and Pacific Biosciences). Other authors do not have competing interests.
Figures















Update of
-
Single-strand mismatch and damage patterns revealed by single-molecule DNA sequencing.bioRxiv [Preprint]. 2023 Feb 19:2023.02.19.526140. doi: 10.1101/2023.02.19.526140. bioRxiv. 2023. Update in: Nature. 2024 Jun;630(8017):752-761. doi: 10.1038/s41586-024-07532-8. PMID: 36824744 Free PMC article. Updated. Preprint.
References
-
- Mustjoki S & Young NS Somatic Mutations in “Benign” Disease. New England Journal of Medicine 384, 2039–2052 (2021). - PubMed
-
- Seplyarskiy VB & Sunyaev S The origin of human mutation in light of genomic data. Nature Reviews Genetics 22, 672–686 (2021). - PubMed
-
- Koh G, Degasperi A, Zou X, Momen S & Nik-Zainal S Mutational signatures: emerging concepts, caveats and clinical applications. Nature Reviews Cancer 21, 619–637 (2021). - PubMed
Extended Data Figures and Methods References
-
- Agarwal A, Gupta S & Sharma R in Andrological Evaluation of Male Infertility: A Laboratory Guide (eds Ashok Agarwal, Sajal Gupta, & Rakesh Sharma) 101–107 (Springer International Publishing, 2016).
-
- Jenkins TG, Liu L, Aston KI & Carrell DT Pre-screening method for somatic cell contamination in human sperm epigenetic studies. Systems Biology in Reproductive Medicine 64, 146–155 (2018). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
