

The venom and telopodal defence systems of the centipede Lithobius forficatus are functionally convergent serial homologues
- PMID: 38867210
- PMCID: PMC11170834
- DOI: 10.1186/s12915-024-01925-x
The venom and telopodal defence systems of the centipede Lithobius forficatus are functionally convergent serial homologues
Abstract
Background: Evolution of novelty is a central theme in evolutionary biology, yet studying the origins of traits with an apparently discontinuous origin remains a major challenge. Venom systems are a well-suited model for the study of this phenomenon because they capture several aspects of novelty across multiple levels of biological complexity. However, while there is some knowledge on the evolution of individual toxins, not much is known about the evolution of venom systems as a whole. One way of shedding light on the evolution of new traits is to investigate less specialised serial homologues, i.e. repeated traits in an organism that share a developmental origin. This approach can be particularly informative in animals with repetitive body segments, such as centipedes.
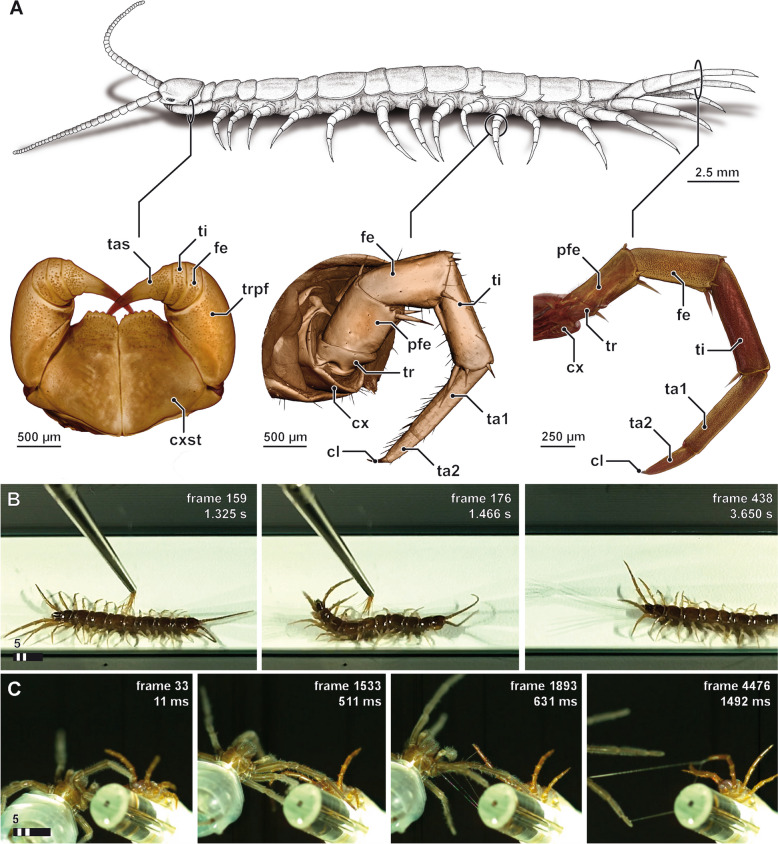
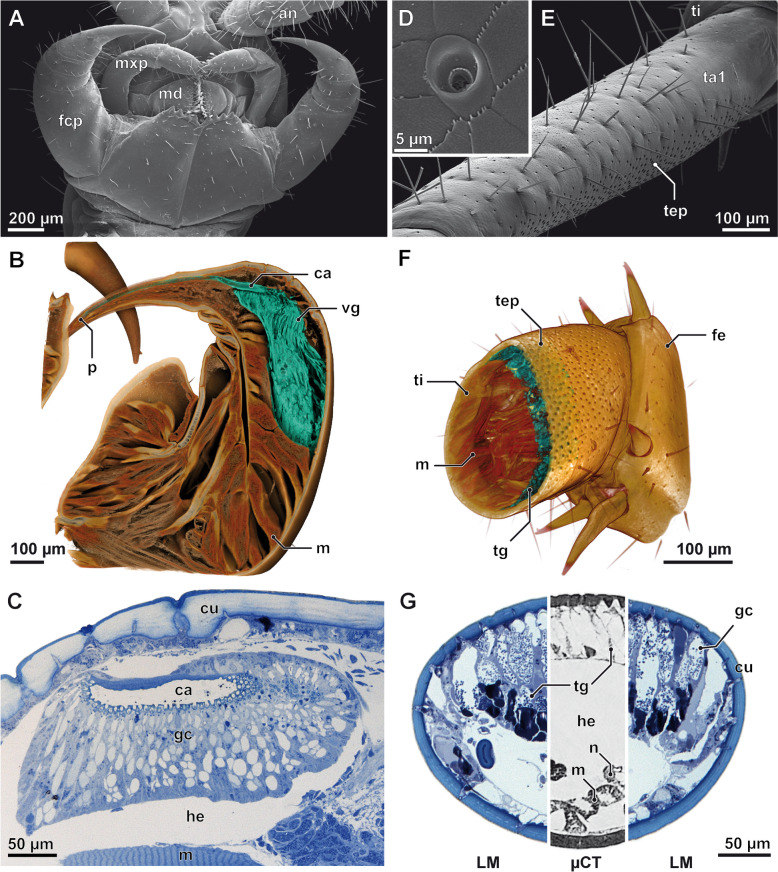
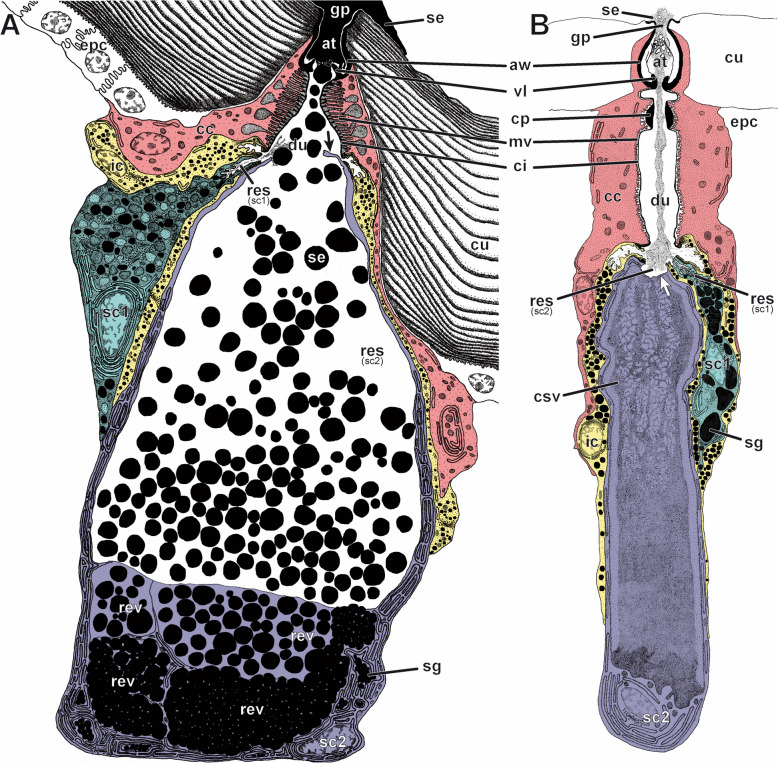
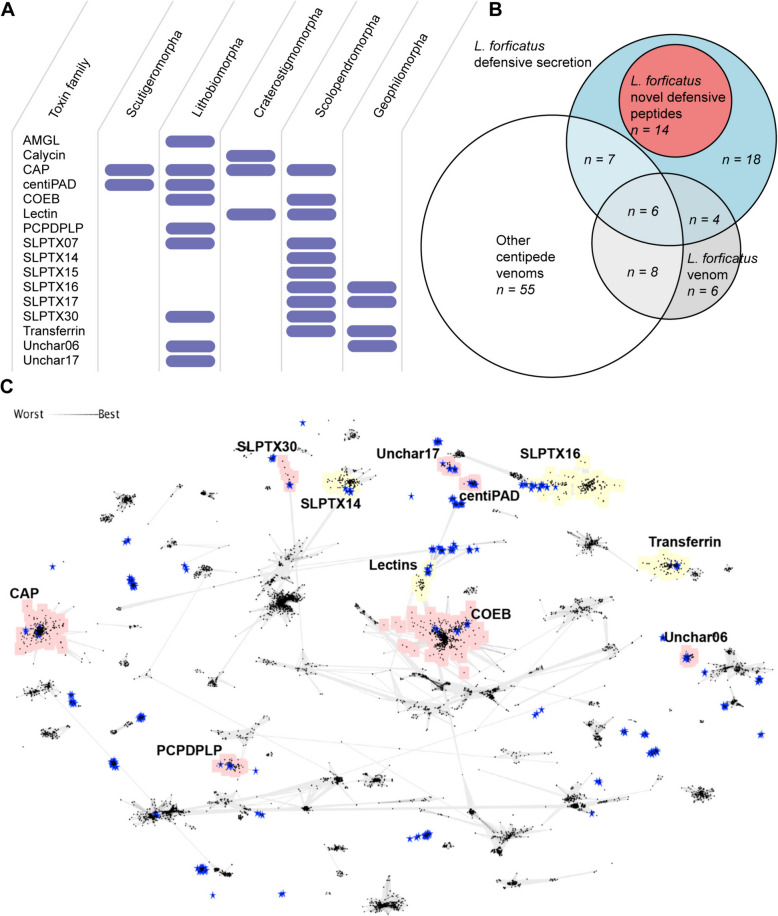
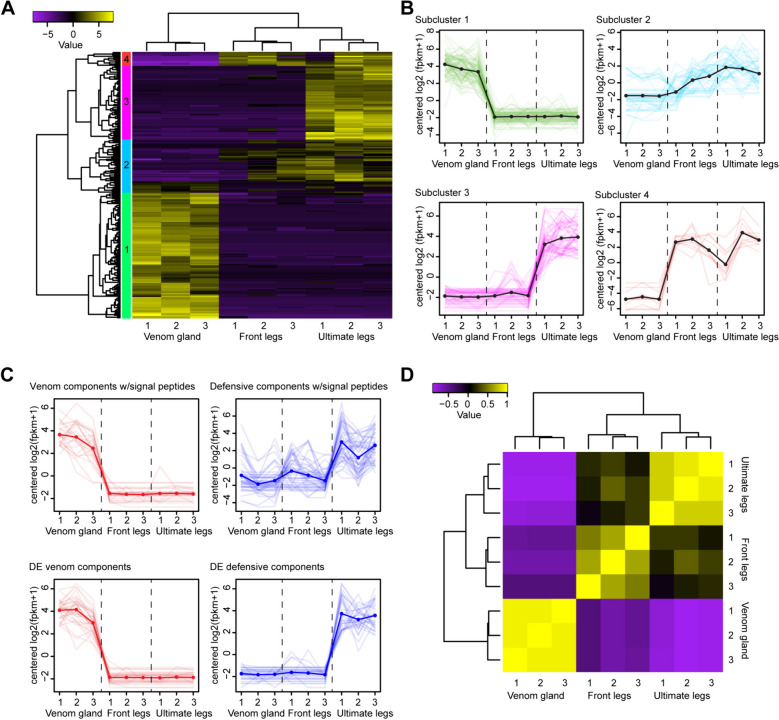
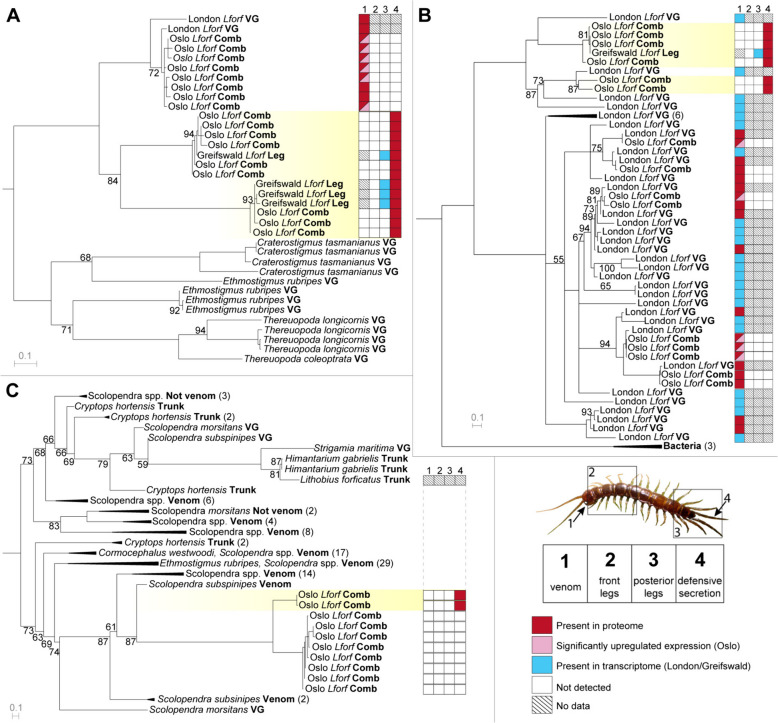
Results: Here, we investigate morphological and biochemical aspects of the defensive telopodal glandular organs borne on the posterior legs of venomous stone centipedes (Lithobiomorpha), using a multimethod approach, including behavioural observations, comparative morphology, proteomics, comparative transcriptomics and molecular phylogenetics. We show that the anterior venom system and posterior telopodal defence system are functionally convergent serial homologues, where one (telopodal defence) represents a model for the putative early evolutionary state of the other (venom). Venom glands and telopodal glandular organs appear to have evolved from the same type of epidermal gland (four-cell recto-canal type) and while the telopodal defensive secretion shares a great degree of compositional overlap with centipede venoms in general, these similarities arose predominantly through convergent recruitment of distantly related toxin-like components. Both systems are composed of elements predisposed to functional innovation across levels of biological complexity that range from proteins to glands, demonstrating clear parallels between molecular and morphological traits in the properties that facilitate the evolution of novelty.
Conclusions: The evolution of the lithobiomorph telopodal defence system provides indirect empirical support for the plausibility of the hypothesised evolutionary origin of the centipede venom system, which occurred through functional innovation and gradual specialisation of existing epidermal glands. Our results thus exemplify how continuous transformation and functional innovation can drive the apparent discontinuous emergence of novelties on higher levels of biological complexity.
Keywords: Arthropoda; Chilopoda; Epidermal exocrine glands; Evolution; Innovation; Myriapoda; Novelty; Telopodal glandular organs; Venom.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Exaptation of an evolutionary constraint enables behavioural control over the composition of secreted venom in a giant centipede.Nat Ecol Evol. 2025 Jan;9(1):73-86. doi: 10.1038/s41559-024-02556-9. Epub 2024 Nov 4. Nat Ecol Evol. 2025. PMID: 39496866
-
Parallel Evolution of Complex Centipede Venoms Revealed by Comparative Proteotranscriptomic Analyses.Mol Biol Evol. 2019 Dec 1;36(12):2748-2763. doi: 10.1093/molbev/msz181. Mol Biol Evol. 2019. PMID: 31396628 Free PMC article.
-
Production and packaging of a biological arsenal: evolution of centipede venoms under morphological constraint.Proc Natl Acad Sci U S A. 2015 Mar 31;112(13):4026-31. doi: 10.1073/pnas.1424068112. Epub 2015 Mar 16. Proc Natl Acad Sci U S A. 2015. PMID: 25775536 Free PMC article.
-
Centipede venoms and their components: resources for potential therapeutic applications.Toxins (Basel). 2015 Nov 17;7(11):4832-51. doi: 10.3390/toxins7114832. Toxins (Basel). 2015. PMID: 26593947 Free PMC article. Review.
-
Centipede venom: recent discoveries and current state of knowledge.Toxins (Basel). 2015 Feb 25;7(3):679-704. doi: 10.3390/toxins7030679. Toxins (Basel). 2015. PMID: 25723324 Free PMC article. Review.
Cited by
-
Placozoan secretory cell types implicated in feeding, innate immunity and regulation of behavior.bioRxiv [Preprint]. 2025 Jan 16:2024.09.18.613768. doi: 10.1101/2024.09.18.613768. bioRxiv. 2025. Update in: PLoS One. 2025 Apr 22;20(4):e0311271. doi: 10.1371/journal.pone.0311271. PMID: 39372748 Free PMC article. Updated. Preprint.
-
Placozoan secretory cell types implicated in feeding, innate immunity and regulation of behavior.PLoS One. 2025 Apr 22;20(4):e0311271. doi: 10.1371/journal.pone.0311271. eCollection 2025. PLoS One. 2025. PMID: 40261937 Free PMC article.
References
-
- Pigliucci M. What, if anything, is an evolutionary novelty? Philos Sci. 2008;75:887–898. doi: 10.1086/594532. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
