

Antibody signatures against viruses and microbiome reflect past and chronic exposures and associate with aging and inflammation
- PMID: 38868191
- PMCID: PMC11167443
- DOI: 10.1016/j.isci.2024.109981
Antibody signatures against viruses and microbiome reflect past and chronic exposures and associate with aging and inflammation
Abstract
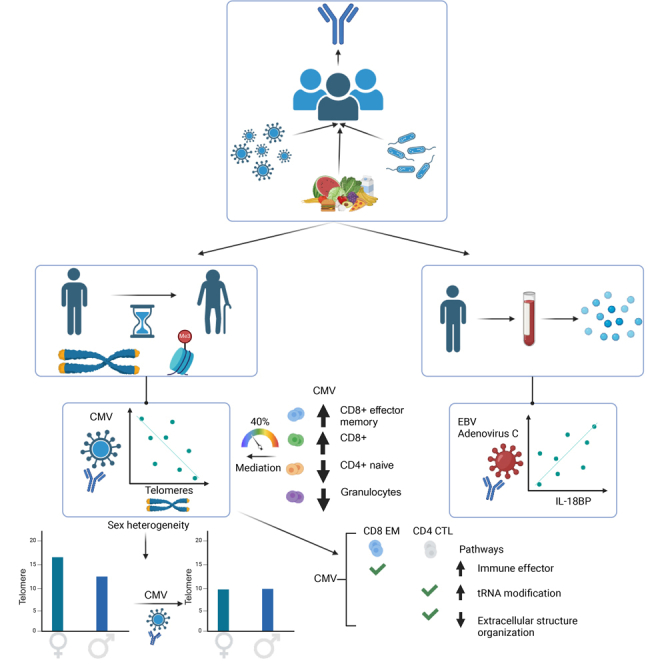
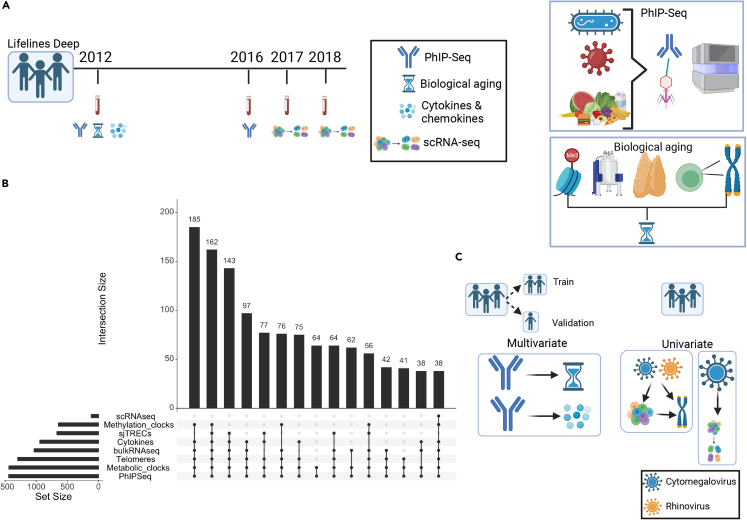
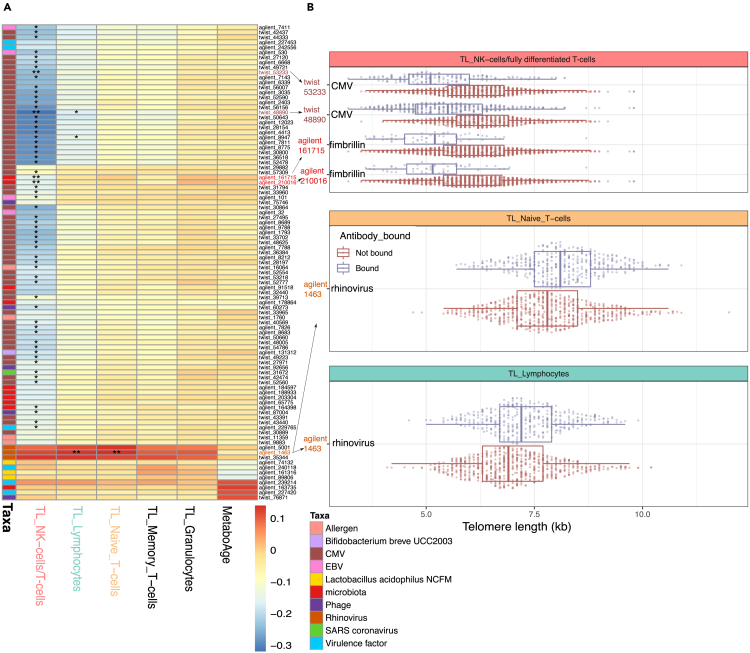
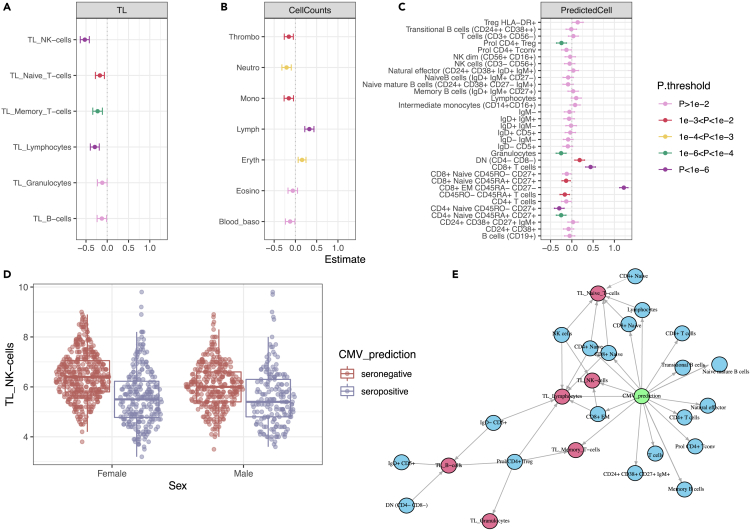
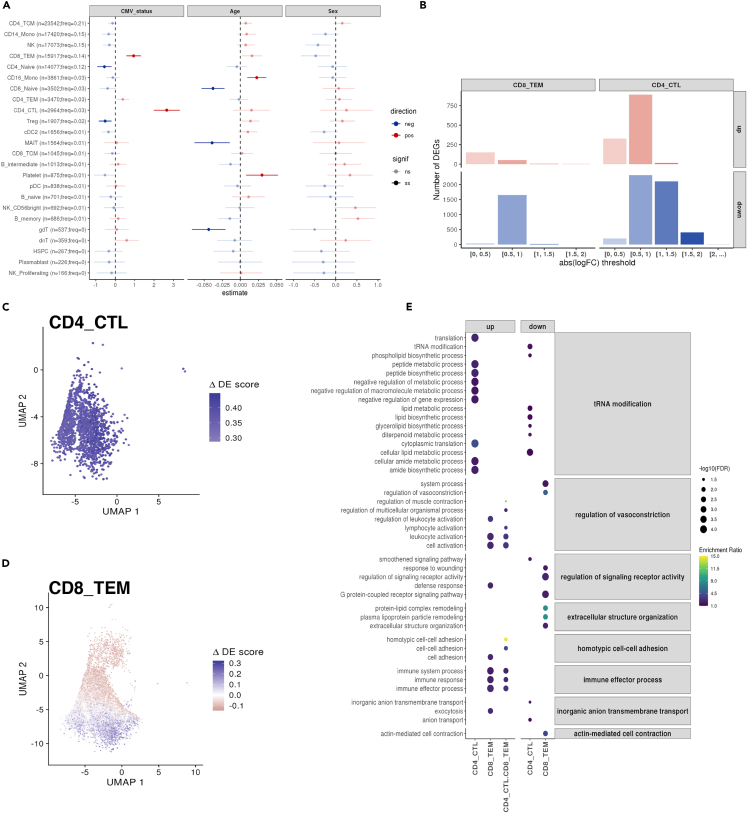
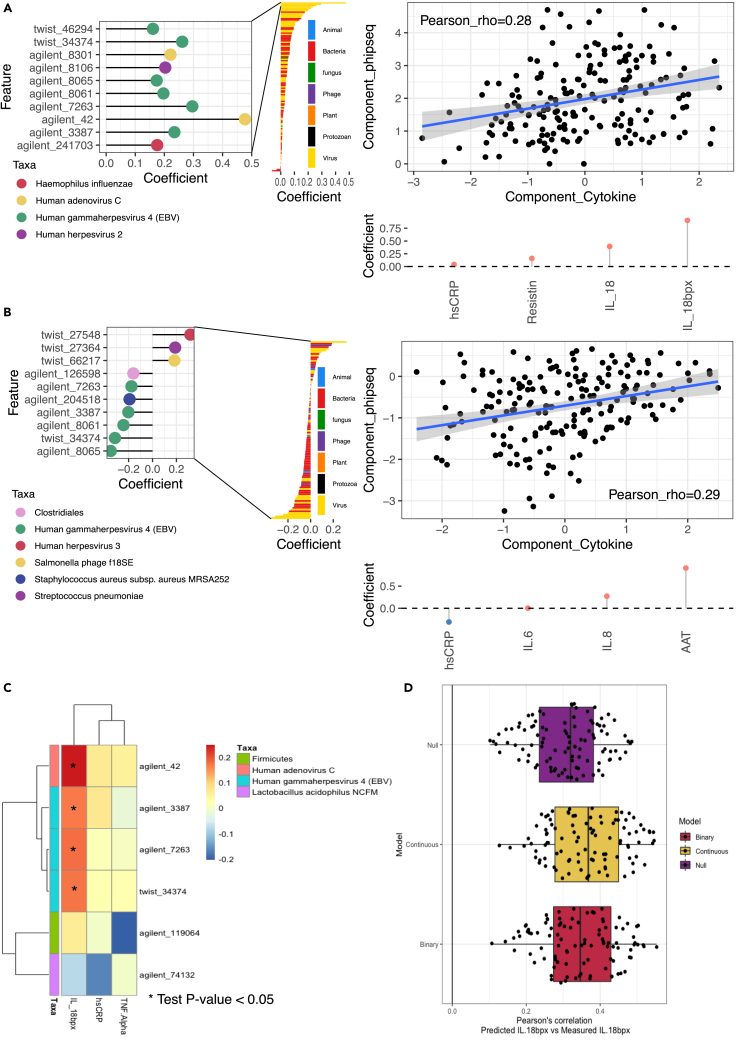
Encounters with pathogens and other molecules can imprint long-lasting effects on our immune system, influencing future physiological outcomes. Given the wide range of microbes to which humans are exposed, their collective impact on health is not fully understood. To explore relations between exposures and biological aging and inflammation, we profiled an antibody-binding repertoire against 2,815 microbial, viral, and environmental peptides in a population cohort of 1,443 participants. Utilizing antibody-binding as a proxy for past exposures, we investigated their impact on biological aging, cell composition, and inflammation. Immune response against cytomegalovirus (CMV), rhinovirus, and gut bacteria relates with telomere length. Single-cell expression measurements identified an effect of CMV infection on the transcriptional landscape of subpopulations of CD8 and CD4 T-cells. This examination of the relationship between microbial exposures and biological aging and inflammation highlights a role for chronic infections (CMV and Epstein-Barr virus) and common pathogens (rhinoviruses and adenovirus C).
Keywords: Genomics; Immunology; Proteomics.
© 2024 The Authors.
Conflict of interest statement
P.L. is a founding shareholder in Repeat Diagnostics, a CLIA certified company specializing in leukocyte telomere length measurements using Flow-FISH, where G.A. is also employed. The other authors declare no competing interests.
Figures
References
-
- Nikolich-Žugich J. The twilight of immunity: emerging concepts in aging of the immune system. Nat. Immunol. 2018;19:10–19. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
