

Manganese porphyrin-based treatment improves fetal-placental development and protects against oxidative damage and NLRP3 inflammasome activation in a rat maternal hypothyroidism model
- PMID: 38870780
- PMCID: PMC11225907
- DOI: 10.1016/j.redox.2024.103238
Manganese porphyrin-based treatment improves fetal-placental development and protects against oxidative damage and NLRP3 inflammasome activation in a rat maternal hypothyroidism model
Abstract
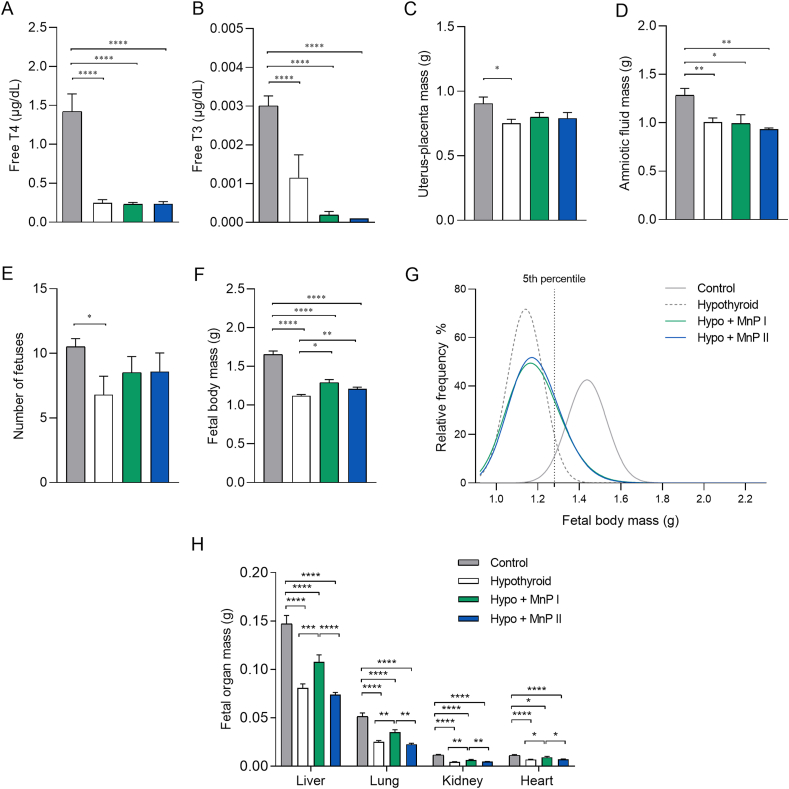
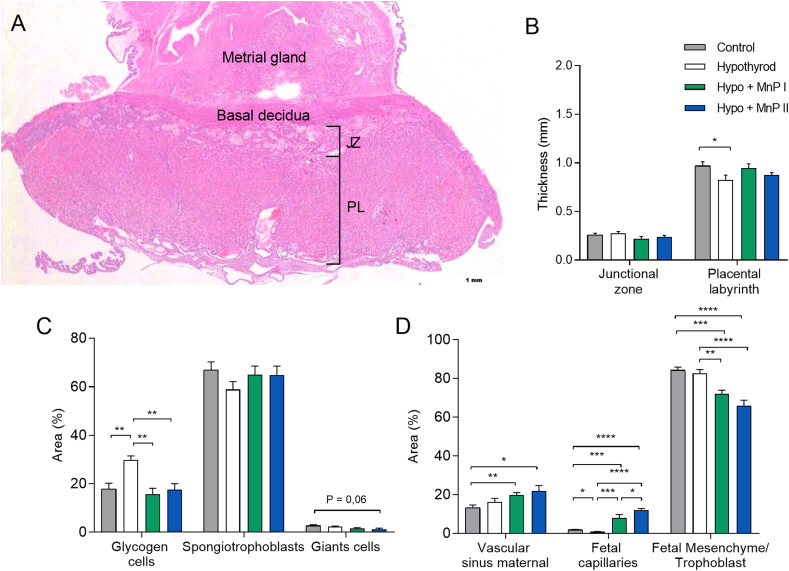
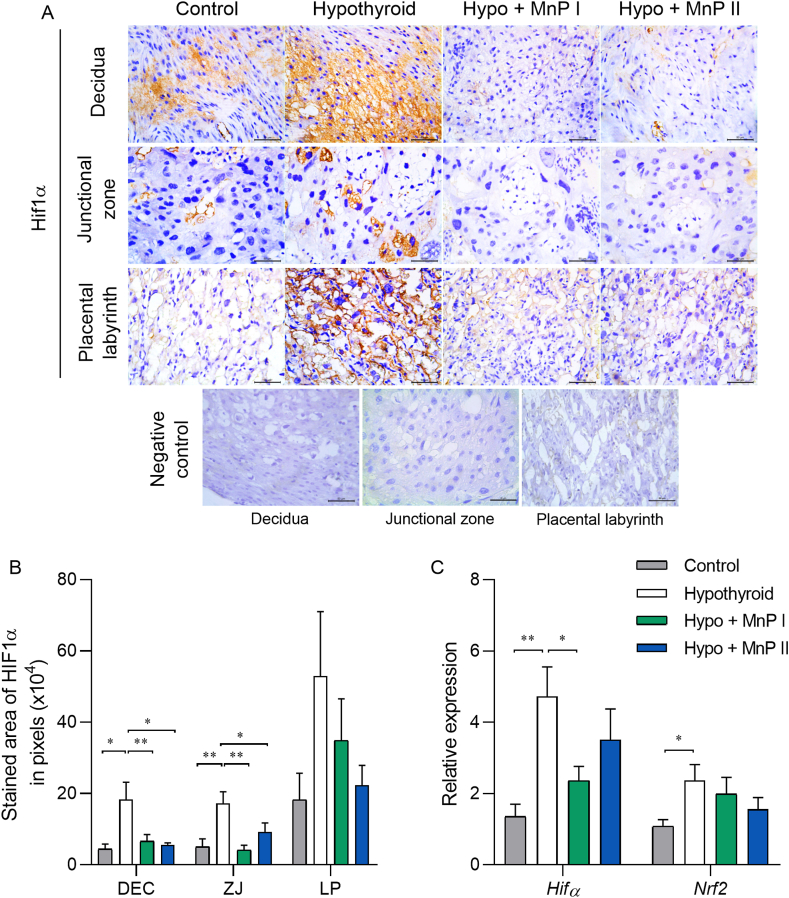
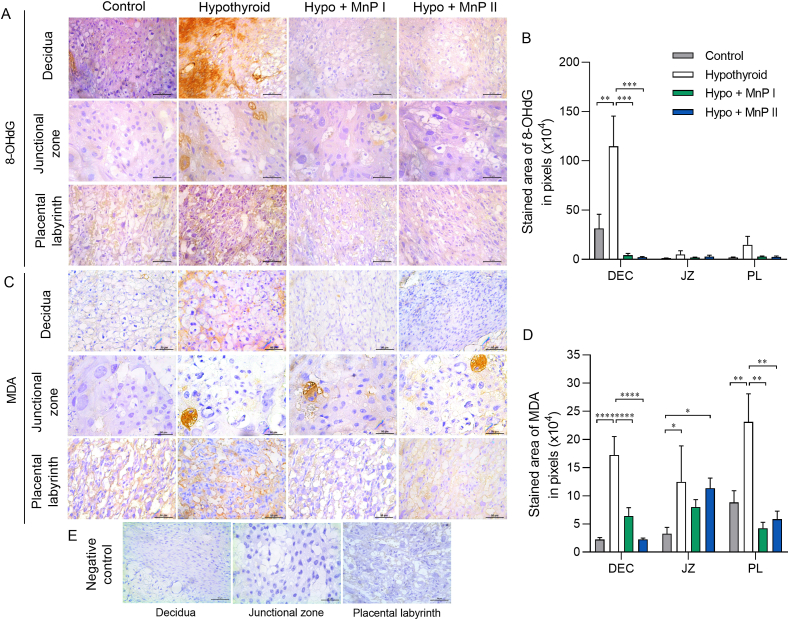
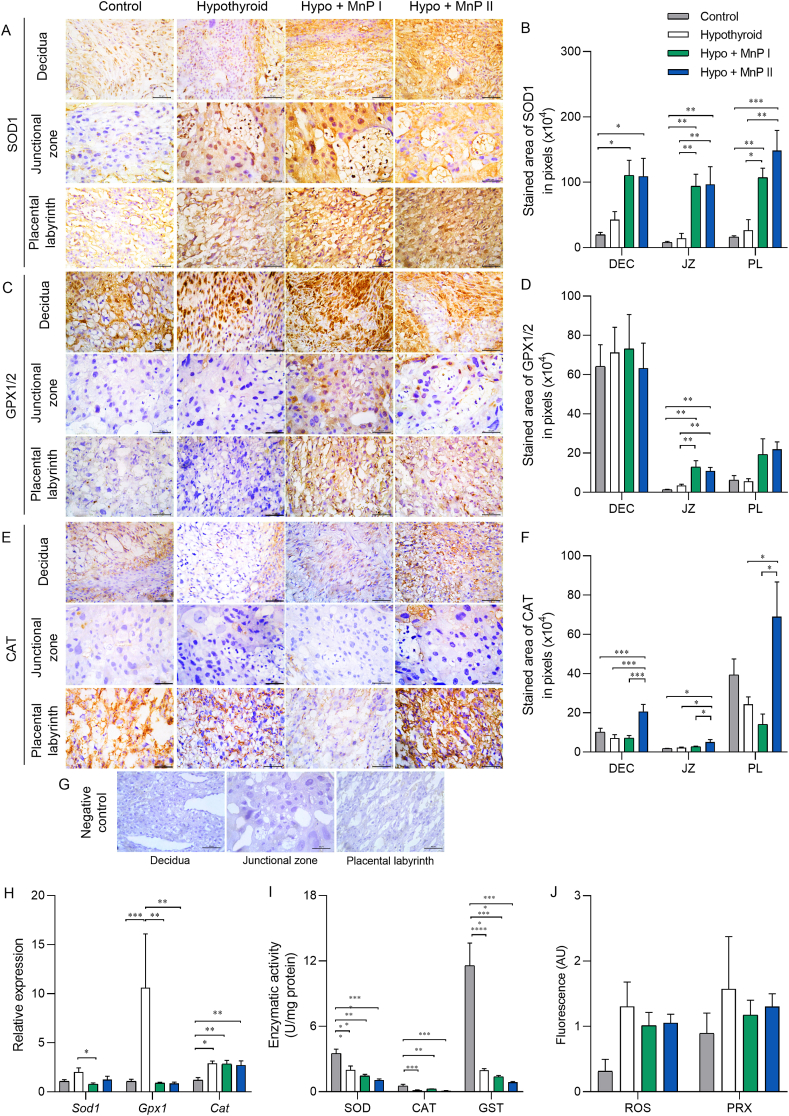
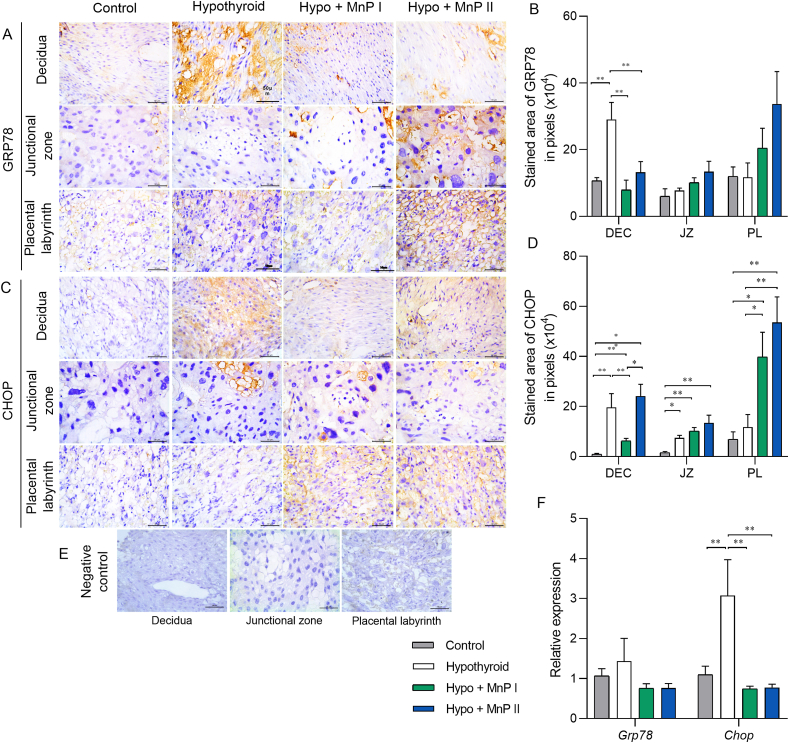
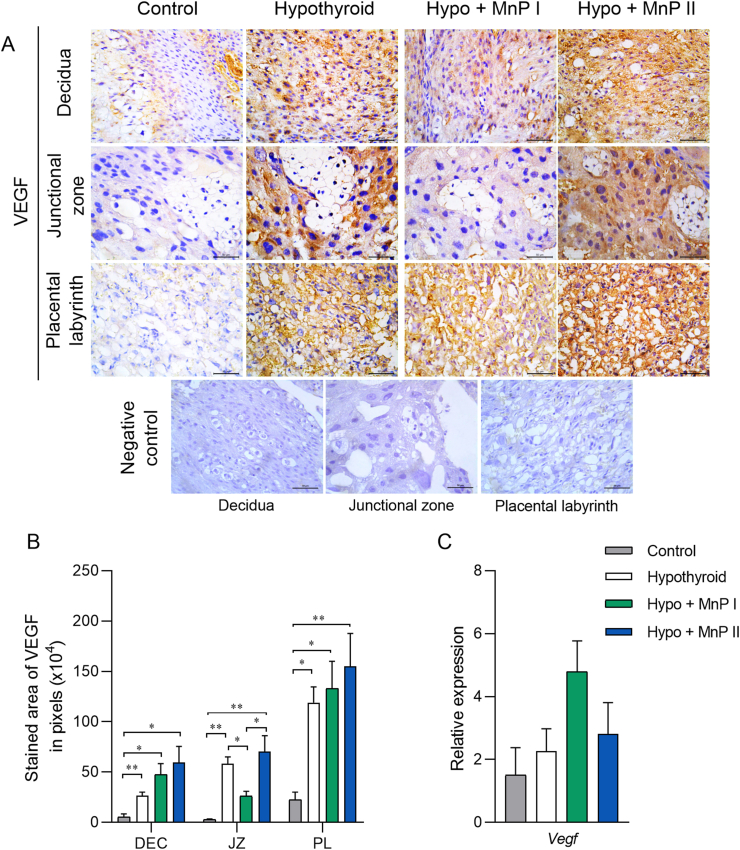
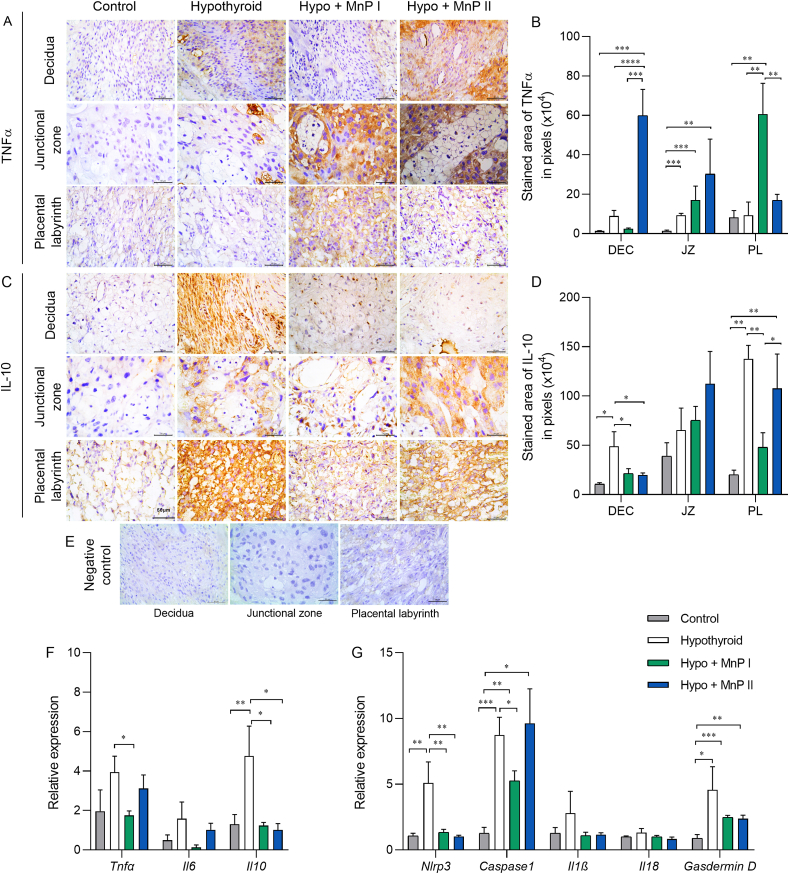
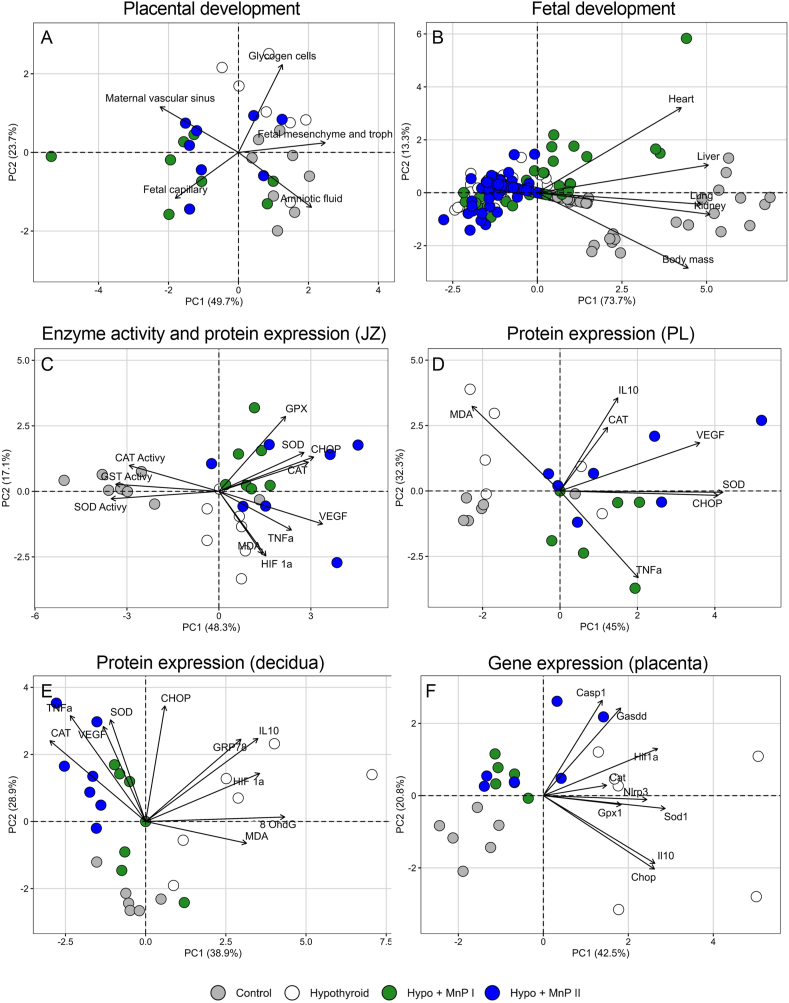
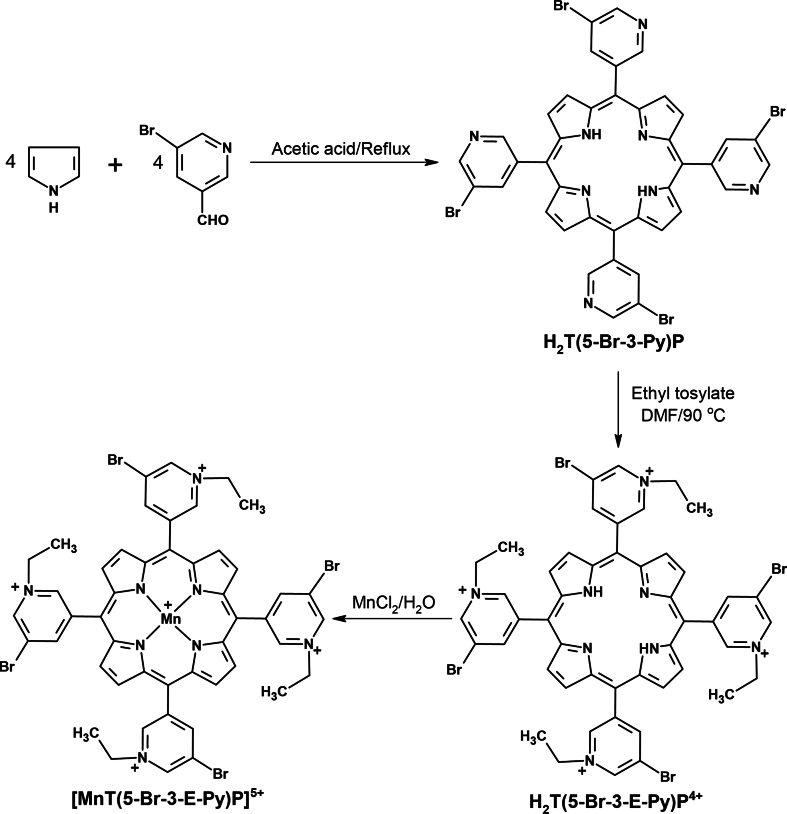
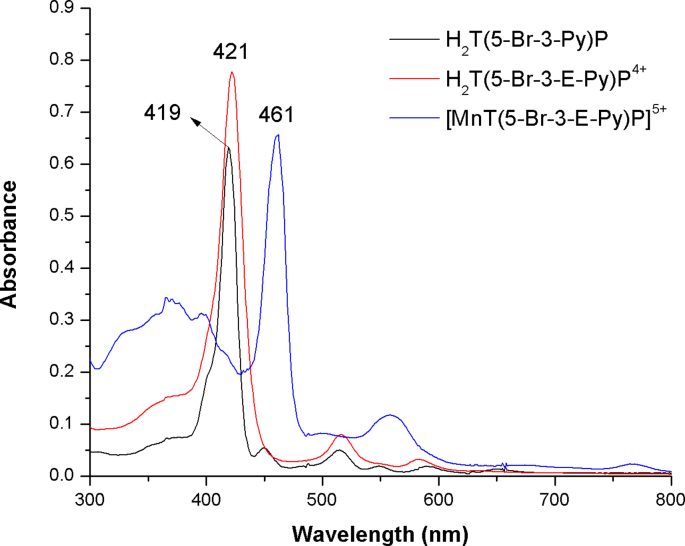
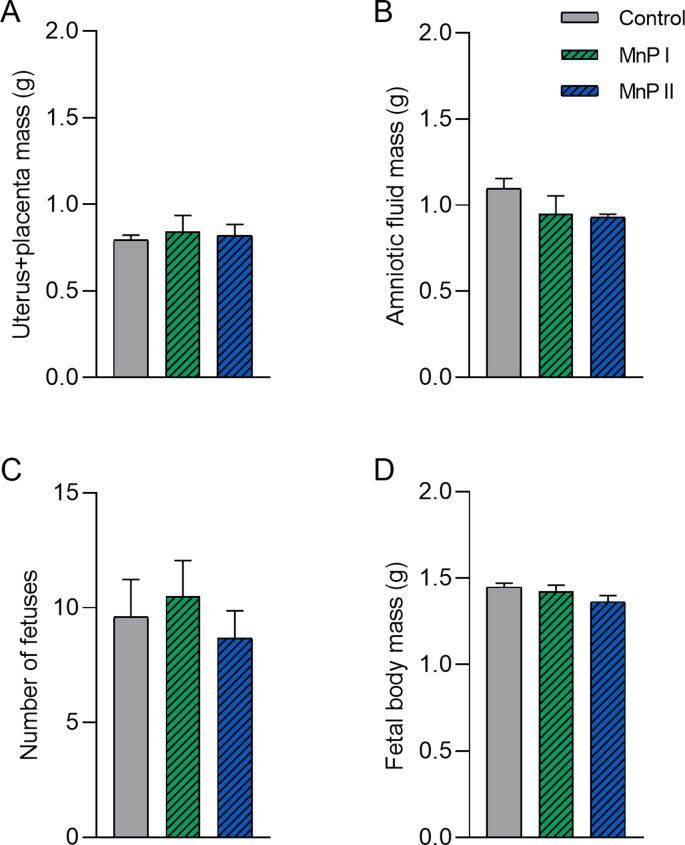
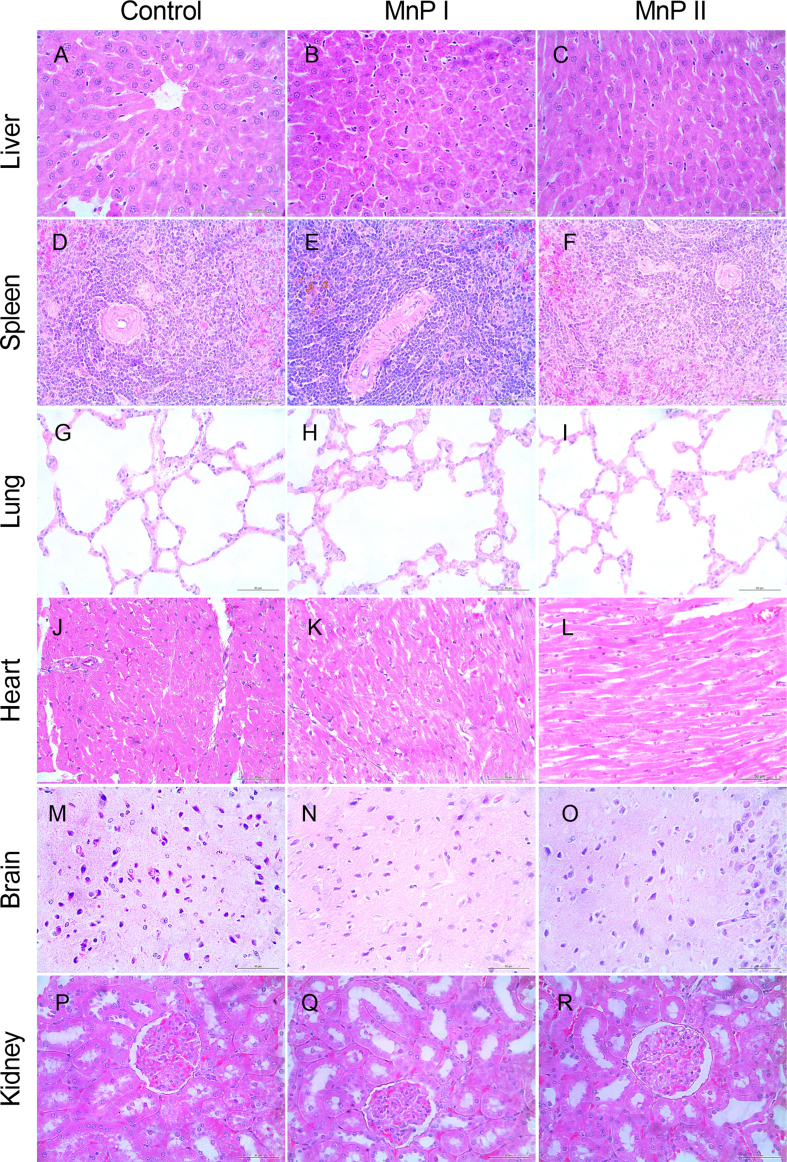
Oxidative stress (OS) and endoplasmic reticulum stress (ERS) are at the genesis of placental disorders observed in preeclampsia, intrauterine growth restriction, and maternal hypothyroidism. In this regard, cationic manganese porphyrins (MnPs) comprise potent redox-active therapeutics of high antioxidant and anti-inflammatory potential, which have not been evaluated in metabolic gestational diseases yet. This study evaluated the therapeutic potential of two MnPs, [MnTE-2-PyP]5+ (MnP I) and [MnT(5-Br-3-E-Py)P]5+ (MnP II), in the fetal-placental dysfunction of hypothyroid rats. Hypothyroidism was induced by administration of 6-Propyl-2-thiouracil (PTU) and treatment with MnPs I and II 0.1 mg/kg/day started on the 8th day of gestation (DG). The fetal and placental development, and protein and/or mRNA expression of antioxidant mediators (SOD1, CAT, GPx1), hypoxia (HIF1α), oxidative damage (8-OHdG, MDA), ERS (GRP78 and CHOP), immunological (TNFα, IL-6, IL-10, IL-1β, IL-18, NLRP3, Caspase1, Gasdermin D) and angiogenic (VEGF) were evaluated in the placenta and decidua on the 18th DG using immunohistochemistry and qPCR. ROS and peroxynitrite (PRX) were quantified by fluorometric assay, while enzyme activities of SOD, GST, and catalase were evaluated by colorimetric assay. MnPs I and II increased fetal body mass in hypothyroid rats, and MnP I increased fetal organ mass. MnPs restored the junctional zone morphology in hypothyroid rats and increased placental vascularization. MnPs blocked the increase of OS and ERS mediators caused by hypothyroidism, showing similar levels of expression of HIFα, 8-OHdG, MDA, Gpx1, GRP78, and Chop to the control. Moreover, MnPs I and/or II increased the protein expression of SOD1, Cat, and GPx1 and restored the expression of IL10, Nlrp3, and Caspase1 in the decidua and/or placenta. However, MnPs did not restore the low placental enzyme activity of SOD, CAT, and GST caused by hypothyroidism, while increased the decidual and placental protein expression of TNFα. The results show that treatment with MnPs improves the fetal-placental development and the placental inflammatory state of hypothyroid rats and protects against oxidative stress and reticular stress caused by hypothyroidism at the maternal-fetal interface.
Keywords: Antioxidant; Inflammasome; Metalloporphyrin; Placenta; Rat; Thyroid.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Maternal hypothyroidism causes oxidative stress and endoplasmic reticulum stress in the maternal-fetal interface of rats.Free Radic Biol Med. 2022 Oct;191:24-39. doi: 10.1016/j.freeradbiomed.2022.08.033. Epub 2022 Aug 26. Free Radic Biol Med. 2022. PMID: 36038036
-
Kisspeptin treatment improves fetal-placental development and blocks placental oxidative damage caused by maternal hypothyroidism in an experimental rat model.Front Endocrinol (Lausanne). 2022 Jul 28;13:908240. doi: 10.3389/fendo.2022.908240. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35966095 Free PMC article.
-
Kisspeptin Suppresses Inflammasome-NLRP3 Activation and Pyroptosis Caused by Hypothyroidism at the Maternal-Fetal Interface of Rats.Int J Mol Sci. 2023 Apr 6;24(7):6820. doi: 10.3390/ijms24076820. Int J Mol Sci. 2023. PMID: 37047793 Free PMC article.
-
Diverse functions of cationic Mn(III) N-substituted pyridylporphyrins, recognized as SOD mimics.Free Radic Biol Med. 2011 Sep 1;51(5):1035-53. doi: 10.1016/j.freeradbiomed.2011.04.046. Epub 2011 May 6. Free Radic Biol Med. 2011. PMID: 21616142 Free PMC article. Review.
-
Crosstalk between ER stress, NLRP3 inflammasome, and inflammation.Appl Microbiol Biotechnol. 2020 Jul;104(14):6129-6140. doi: 10.1007/s00253-020-10614-y. Epub 2020 May 24. Appl Microbiol Biotechnol. 2020. PMID: 32447438 Review.
Cited by
-
Endoplasmic reticulum stress: an emerging therapeutic target for preeclampsia†.Biol Reprod. 2025 Jul 13;113(1):19-33. doi: 10.1093/biolre/ioaf071. Biol Reprod. 2025. PMID: 40156296 Free PMC article. Review.
-
Protective role of mitophagy on microglia-mediated neuroinflammatory injury through mtDNA-STING signaling in manganese-induced parkinsonism.J Neuroinflammation. 2025 Feb 28;22(1):55. doi: 10.1186/s12974-025-03396-5. J Neuroinflammation. 2025. PMID: 40022162 Free PMC article.
-
Manganese Porphyrin Treatment Improves Redox Status Caused by Acute Compressive Spinal Cord Trauma.Antioxidants (Basel). 2025 May 14;14(5):587. doi: 10.3390/antiox14050587. Antioxidants (Basel). 2025. PMID: 40427469 Free PMC article.
-
Comparison of Endoplasmic Reticulum Stress and Pyroptosis Induced by Pathogenic Calcium Oxalate Monohydrate and Physiologic Calcium Oxalate Dihydrate Crystals in HK-2 Cells: Insights into Kidney Stone Formation.Cells. 2024 Dec 15;13(24):2070. doi: 10.3390/cells13242070. Cells. 2024. PMID: 39768161 Free PMC article.
References
-
- Yung H.W., Calabrese S., Hynx D., Hemmings B.A., Cetin I., Charnock-Jones D.S., Burton G.J. Evidence of placental Translation inhibition and endoplasmic reticulum stress in the etiology of human intrauterine growth restriction. Am. J. Pathol. 2008;173:451–462. doi: 10.2353/AJPATH.2008.071193. - DOI - PMC - PubMed
-
- Burton G.J., Yung H.W., Cindrova-Davies T., Charnock-Jones D.S. Placental endoplasmic reticulum stress and oxidative stress in the pathophysiology of unexplained intrauterine growth restriction and early onset preeclampsia. Placenta. 2009;30:43. doi: 10.1016/J.PLACENTA.2008.11.003. - DOI - PMC - PubMed
-
- Folli F., Corradi D., Fanti P., Davalli A., Paez A., Giaccari A., Perego C., Muscogiuri G. The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro- and macrovascular complications: avenues for a mechanistic-based therapeutic approach. Curr. Diabetes Rev. 2012;7:313–324. doi: 10.2174/157339911797415585. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
