

Correction of age-associated defects in dendritic cells enables CD4+ T cells to eradicate tumors
- PMID: 38870946
- PMCID: PMC11283364
- DOI: 10.1016/j.cell.2024.05.026
Correction of age-associated defects in dendritic cells enables CD4+ T cells to eradicate tumors
Abstract
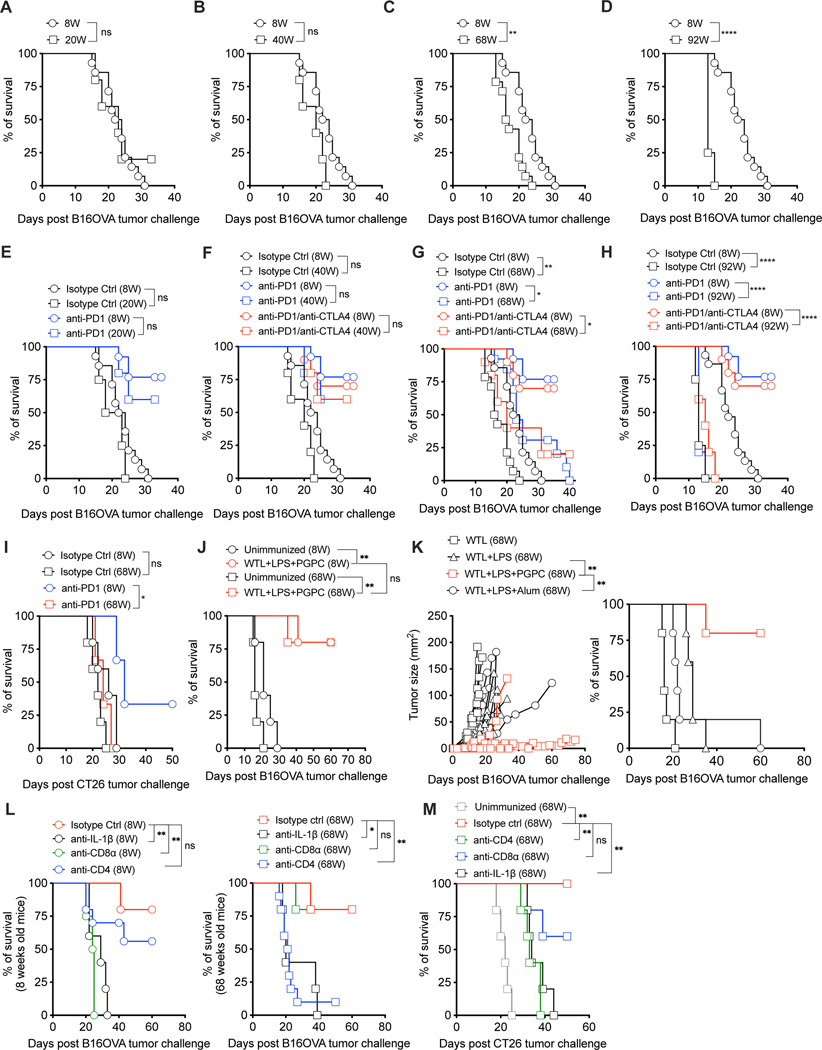
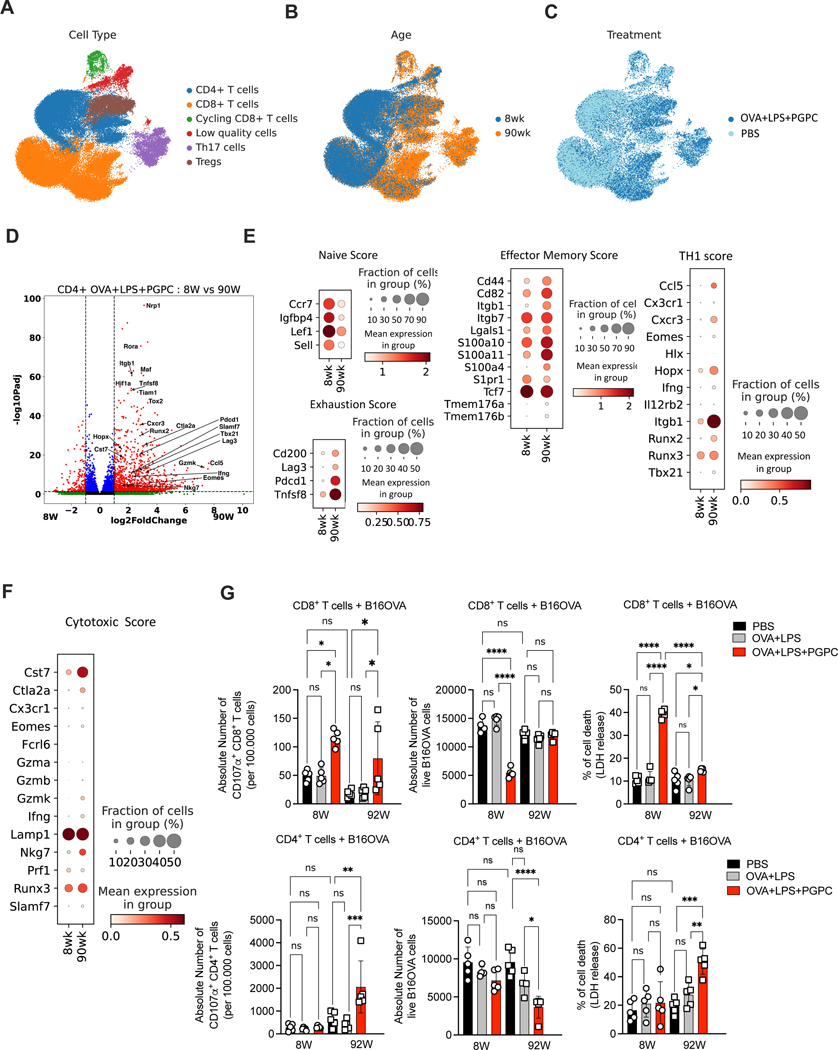
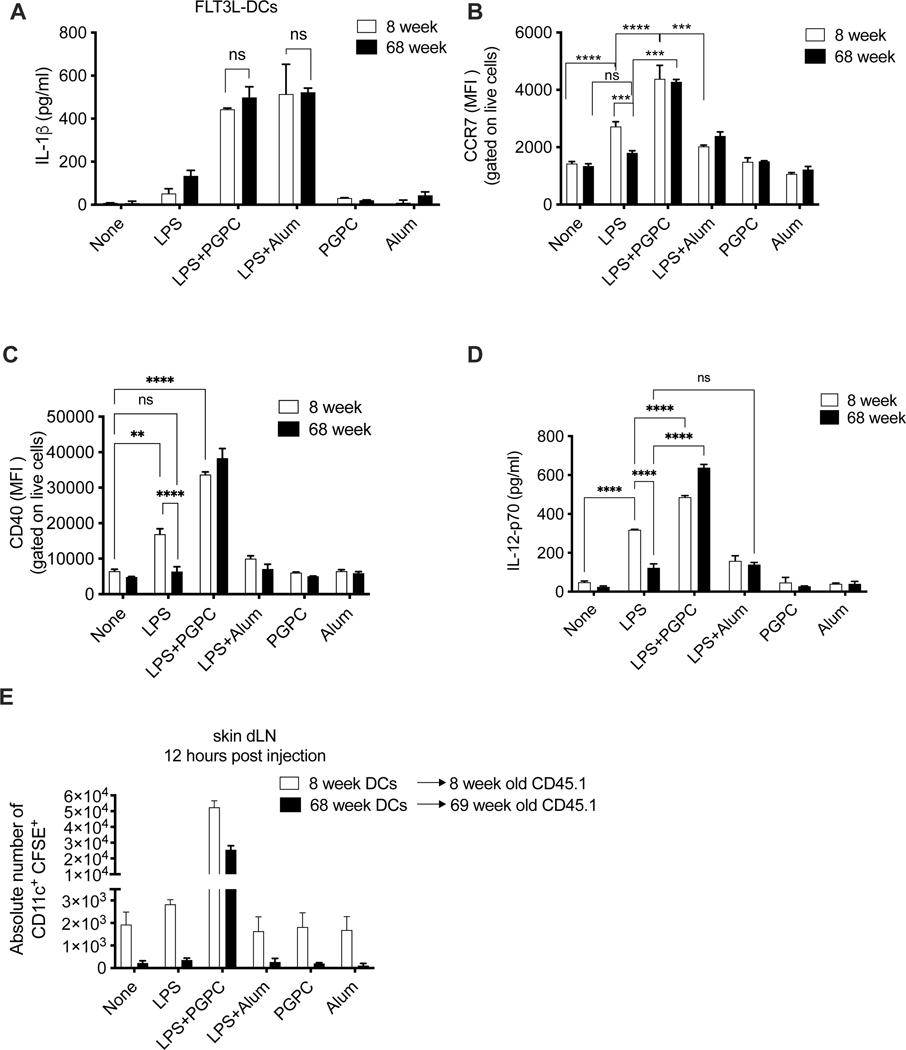
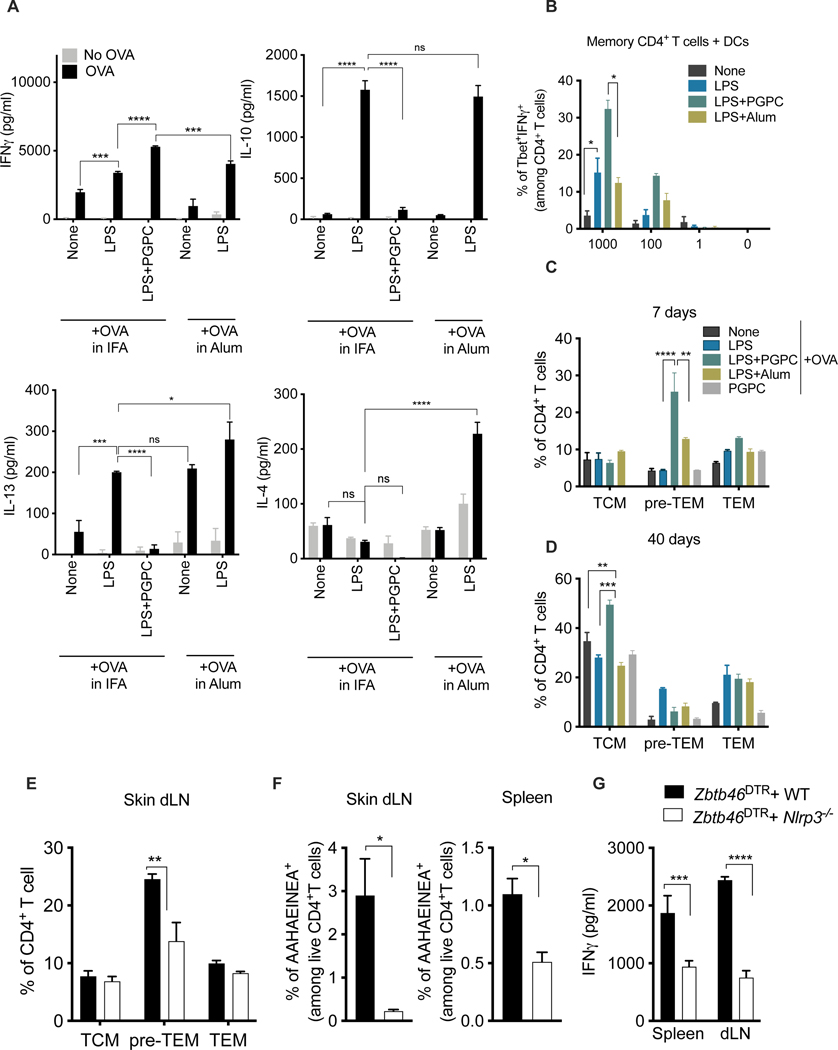
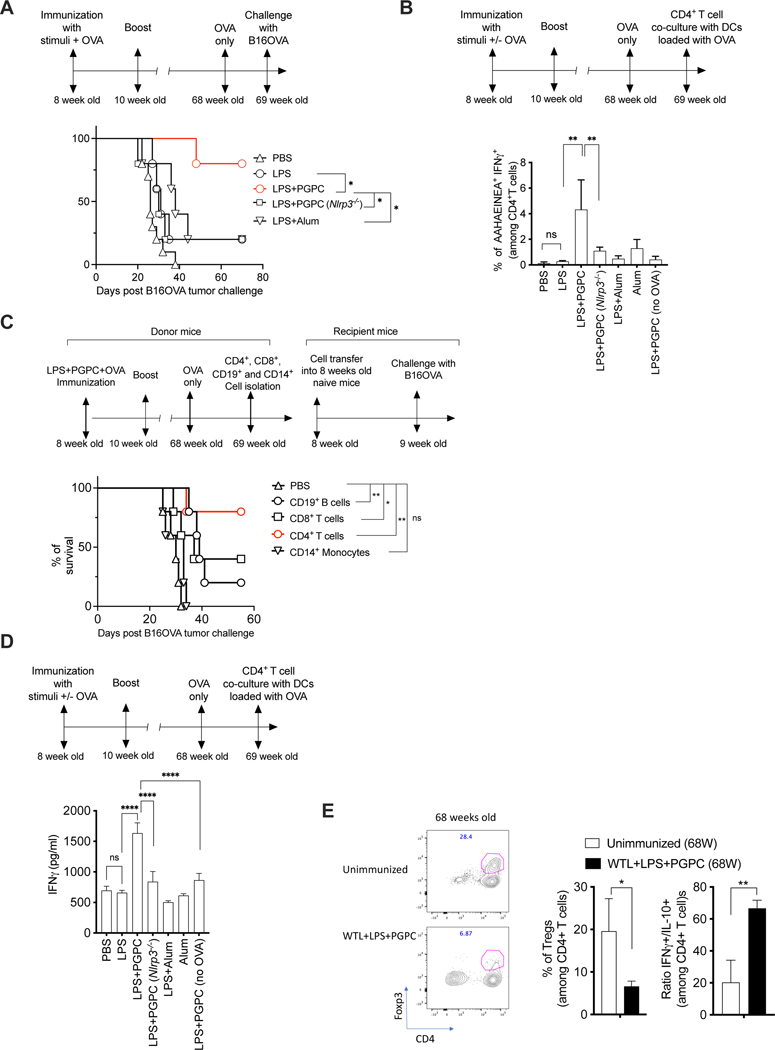
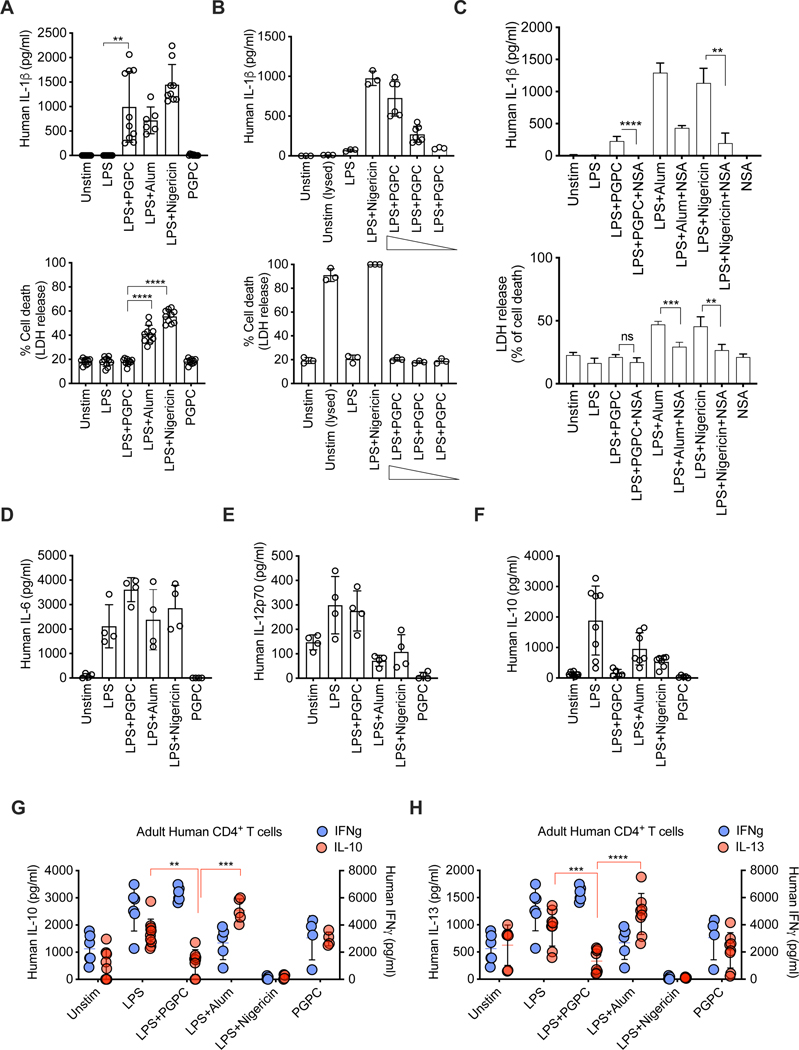
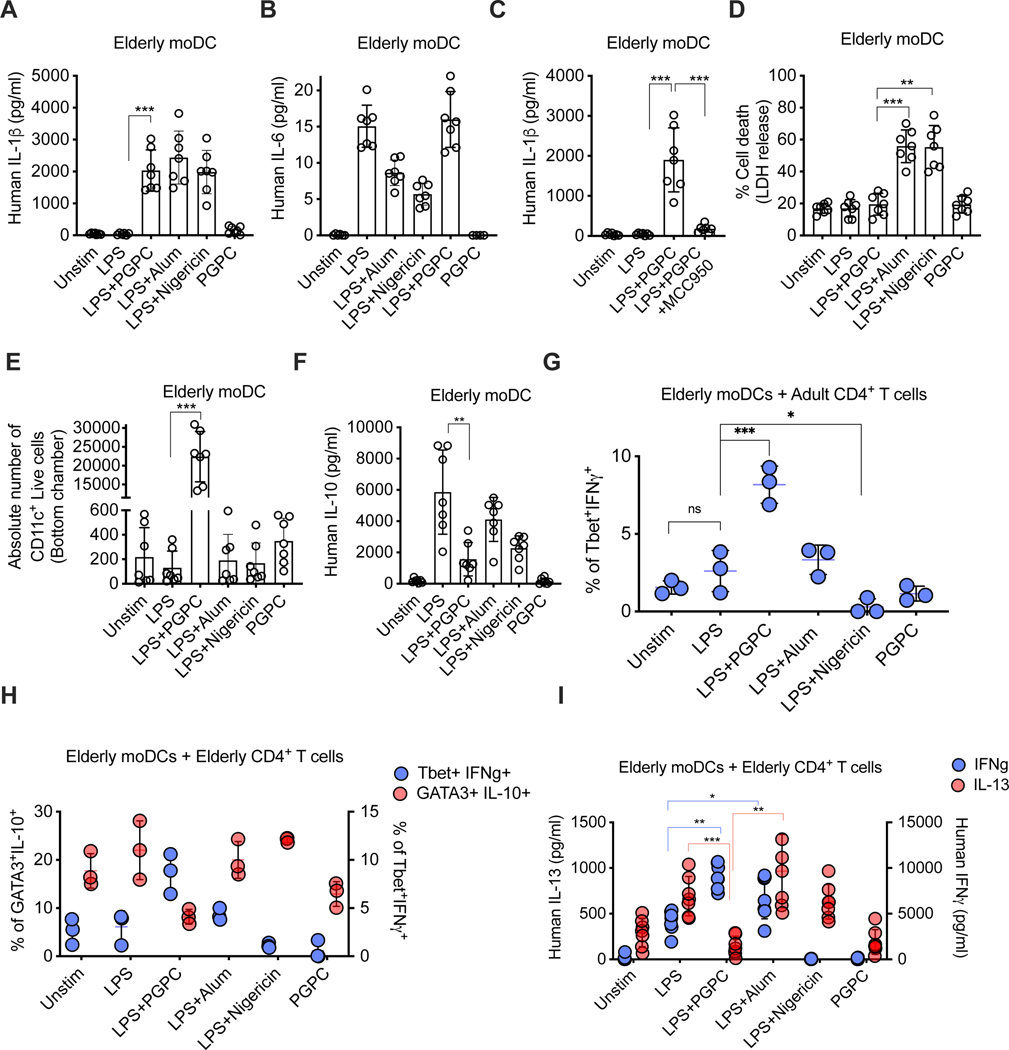
Defective host defenses later in life are associated with changes in immune cell activities, suggesting that age-specific considerations are needed in immunotherapy approaches. In this study, we found that PD-1 and CTLA4-based cancer immunotherapies are unable to eradicate tumors in elderly mice. This defect in anti-tumor activity correlated with two known age-associated immune defects: diminished abundance of systemic naive CD8+ T cells and weak migratory activities of dendritic cells (DCs). We identified a vaccine adjuvant, referred to as a DC hyperactivator, which corrects DC migratory defects in the elderly. Vaccines containing tumor antigens and DC hyperactivators induced T helper type 1 (TH1) CD4+ T cells with cytolytic activity that drive anti-tumor immunity in elderly mice. When administered early in life, DC hyperactivators were the only adjuvant identified that elicited anti-tumor CD4+ T cells that persisted into old age. These results raise the possibility of correcting age-associated immune defects through DC manipulation.
Keywords: PGPC; T cells; cancer; checkpoint inhibitor therapy; dendritic cells; elderly; hyperactivation; immunotherapy; innate immunity; tumor immunity; vaccine.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.C.K. consults and holds equity in Corner Therapeutics, Larkspur Biosciences, MindImmune Therapeutics, and Neumora Therapeutics. R.J.X. is a co-founder of Celsius and Jnana Therapeutics. None of these relationships impacted this study.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
