

The mitochondrial TSPO ligand Atriol mitigates metabolic-associated steatohepatitis by downregulating CXCL1
- PMID: 38871077
- PMCID: PMC11374472
- DOI: 10.1016/j.metabol.2024.155942
The mitochondrial TSPO ligand Atriol mitigates metabolic-associated steatohepatitis by downregulating CXCL1
Abstract
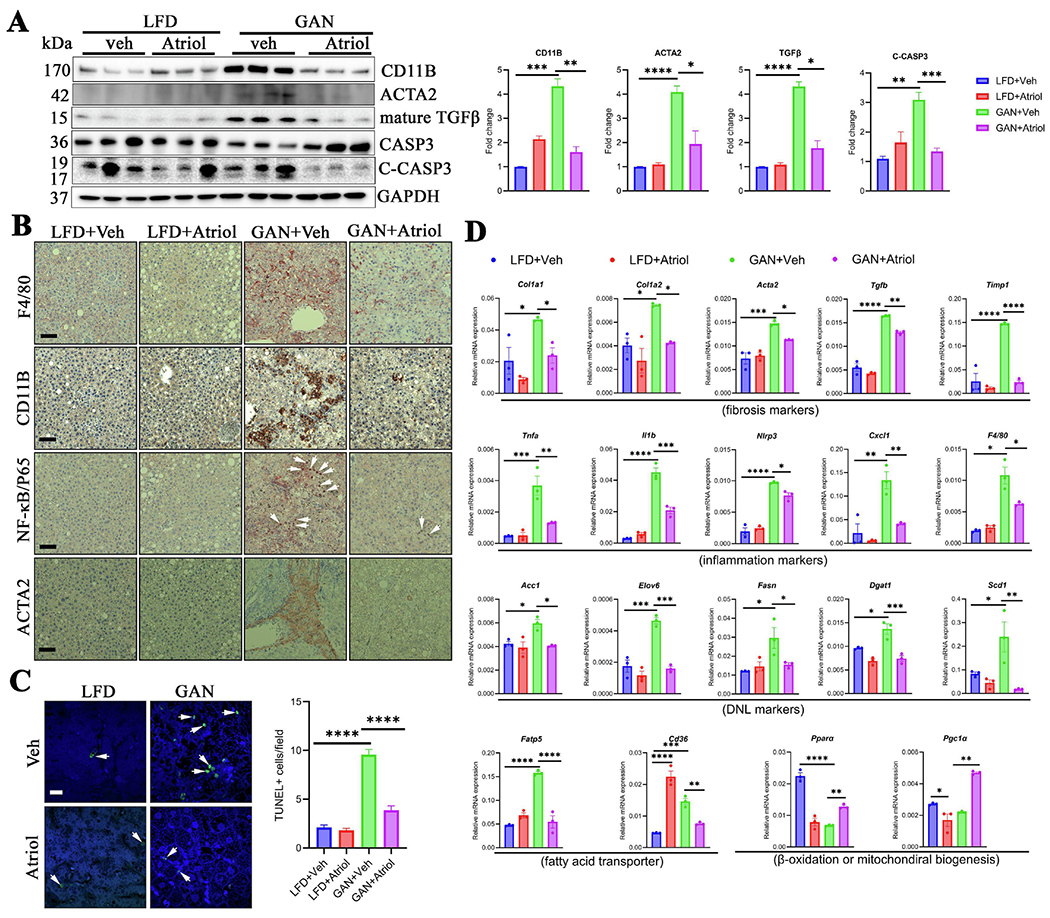
Background and aims: The mitochondrial translocator protein (TSPO, 18 kDa) is pivotal in binding cholesterol and facilitating its transfer from the outer to the inner mitochondrial membrane. Atriol is a TSPO ligand disrupting cholesterol binding by targeting the cholesterol-recognition amino acid consensus domain. Prior research has shown that TSPO deficiency improved metabolic-associated steatohepatitis (MASH). We hypothesized that Atriol may have the potential to alleviate MASH.
Methods and results: In vitro cell culture studies revealed that Atriol treatment effectively mitigated MASH by restoring mitochondrial function, inhibiting the NF-κB signaling pathway, and reducing hepatic stellate cell (HSC) activation. SD male rats were fed a GAN diet for 10 months to induce MASH. During the final two weeks of feeding, rats received intraperitoneal Atriol administration daily. Atriol treatment significantly ameliorated MASH by reducing lipid accumulation, diminishing hepatic lobular inflammation and fibrosis, decreasing cell death, and inhibiting excessive bile acid synthesis. Moreover, Atriol restored mitochondrial function in primary hepatocytes isolated from MASH rats. In search of the mechanism(s) governing these effects, we found that Atriol downregulated the proinflammatory chemokine CXCL1 through the NF-κB signaling pathway or via myeloperoxidase (MPO) in HSCs and Kupffer cells. Additionally, in vitro, studies further suggested that CXCL1 treatment induced dysfunctional mitochondria, inflammation, HSCs activation, and macrophage migration, whereas Atriol countered these effects. Finally, the mitigating effects of Atriol on MASH were reproduced by pharmacological inhibition of NF-κB or MPO and neutralization of CXCL1.
Conclusion: Atriol ameliorates MASH both in vitro and in vivo, demonstrating its potential therapeutic benefits in managing MASH.
Keywords: ADME (Absorption, Distribution, Metabolism, Excretion); Crown-like structures; Fibrosis; Mitochondrial function; Myeloperoxidase; Pharmacokinetic profile; Proinflammation.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of competing interest Go Sugahara is an employee of PhoenixBio, Co., Ltd., Higashi-Hiroshima, Hiroshima, Japan.
Figures






References
-
- Younossi Z, Stepanova M, Ong JP, Jacobson IM, Bugianesi E, Duseja A, Eguchi Y, Wong VW, Negro F, Yilmaz Y, et al. (2019). Nonalcoholic steatohepatitis is the fastest growing cause of hepatocellular carcinoma in liver transplant candidates. Clin Gastroenterol Hepatol 17, 748–755 e743. 10.1016/j.cgh.2018.05.057. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
