

Deciphering vimentin assembly: Bridging theoretical models and experimental approaches
- PMID: 38871297
- PMCID: PMC11267000
- DOI: 10.1016/j.mocell.2024.100080
Deciphering vimentin assembly: Bridging theoretical models and experimental approaches
Abstract
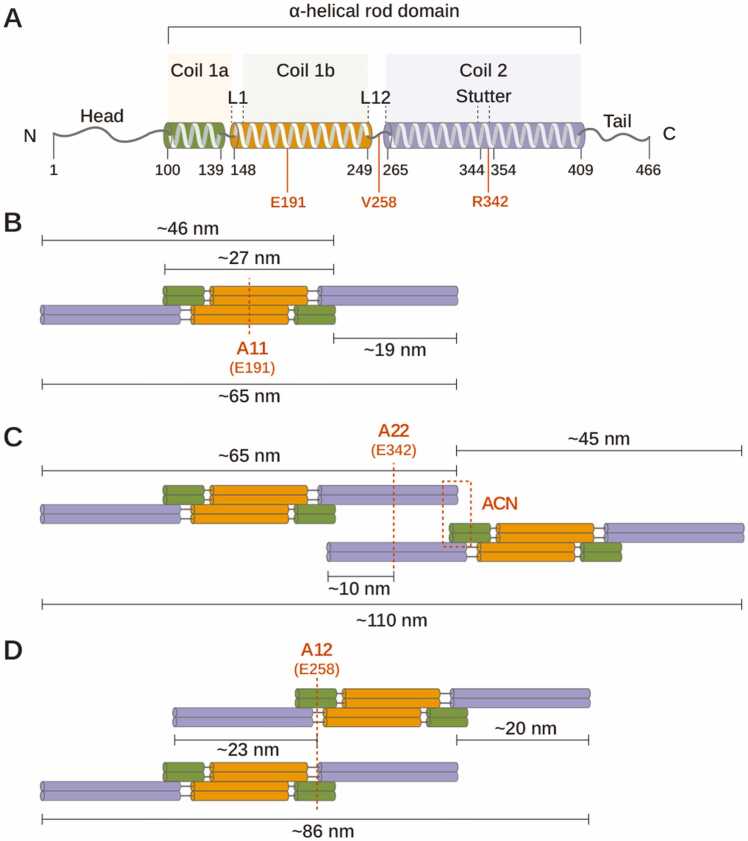
The intricate assembly process of vimentin intermediate filaments (IFs), key components of the eukaryotic cytoskeleton, has yet to be elucidated. In this work, we investigated the transition from soluble tetrameric vimentin units to mature 11-nm tubular filaments, addressing a significant gap in the understanding of IF assembly. Through a combination of theoretical modeling and analysis of experimental data, we propose a novel assembly sequence, emphasizing the role of helical turns and gap filling by soluble tetramers. Our findings shed light on the unique structural dynamics of vimentin and suggest broader implications for the general principles of IF formation.
Keywords: Cytoplasmic intermediate filament; Structural assembly; Theoretical modeling; Tube-like filament; Vimentin.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interests The author is an Editorial Board Member/Editor-in-Chief/Associate Editor/Guest Editor for [Molecules and Cells] and was not involved in the editorial review or the decision to publish this article.
Figures




Similar articles
-
Near-UV circular dichroism reveals structural transitions of vimentin subunits during intermediate filament assembly.J Mol Biol. 2009 Feb 20;386(2):544-53. doi: 10.1016/j.jmb.2008.12.053. Epub 2008 Dec 30. J Mol Biol. 2009. PMID: 19136013
-
Assembly Kinetics of Vimentin Tetramers to Unit-Length Filaments: A Stopped-Flow Study.Biophys J. 2018 May 22;114(10):2408-2418. doi: 10.1016/j.bpj.2018.04.032. Epub 2018 May 10. Biophys J. 2018. PMID: 29754715 Free PMC article.
-
Structural Dynamics of the Vimentin Coiled-coil Contact Regions Involved in Filament Assembly as Revealed by Hydrogen-Deuterium Exchange.J Biol Chem. 2016 Nov 25;291(48):24931-24950. doi: 10.1074/jbc.M116.748145. Epub 2016 Sep 30. J Biol Chem. 2016. PMID: 27694444 Free PMC article.
-
Structural determinants of intermediate filament mechanics.Curr Opin Cell Biol. 2024 Aug;89:102375. doi: 10.1016/j.ceb.2024.102375. Epub 2024 Jun 7. Curr Opin Cell Biol. 2024. PMID: 38850681 Review.
-
Unique Role of Vimentin in the Intermediate Filament Proteins Family.Biochemistry (Mosc). 2024 Apr;89(4):726-736. doi: 10.1134/S0006297924040114. Biochemistry (Mosc). 2024. PMID: 38831508 Review.
References
-
- Aziz A., Hess J.F., Budamagunta M.S., Voss J.C., Kuzin A.P., Huang Y.P.J., Xiao R., Montelione G.T., FitzGerald P.G., Hunt J.F. The structure of vimentin linker 1 and rod 1b domains characterized by site-directed spin-labeling electron paramagnetic resonance (SDSL-EPR) and X-ray crystallography. J. Biol. Chem. 2012;287:28349–28361. - PMC - PubMed
-
- Bott C.J., Winckler B. Intermediate filaments in developing neurons: beyond structure. Cytoskeleton. 2020;77:110–128. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
