

Histone serotonylation in dorsal raphe nucleus contributes to stress- and antidepressant-mediated gene expression and behavior
- PMID: 38871707
- PMCID: PMC11176395
- DOI: 10.1038/s41467-024-49336-4
Histone serotonylation in dorsal raphe nucleus contributes to stress- and antidepressant-mediated gene expression and behavior
Abstract
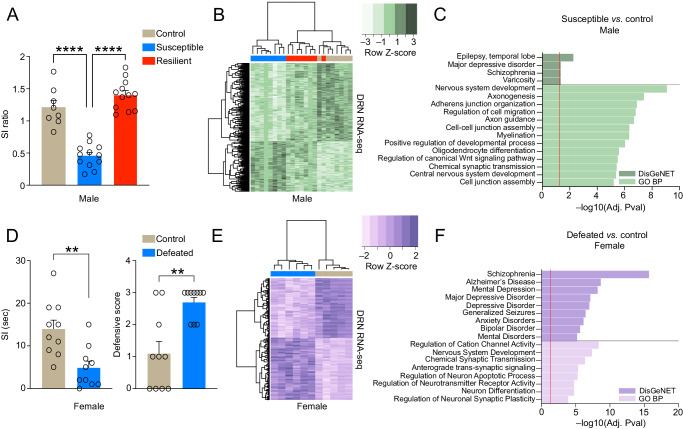
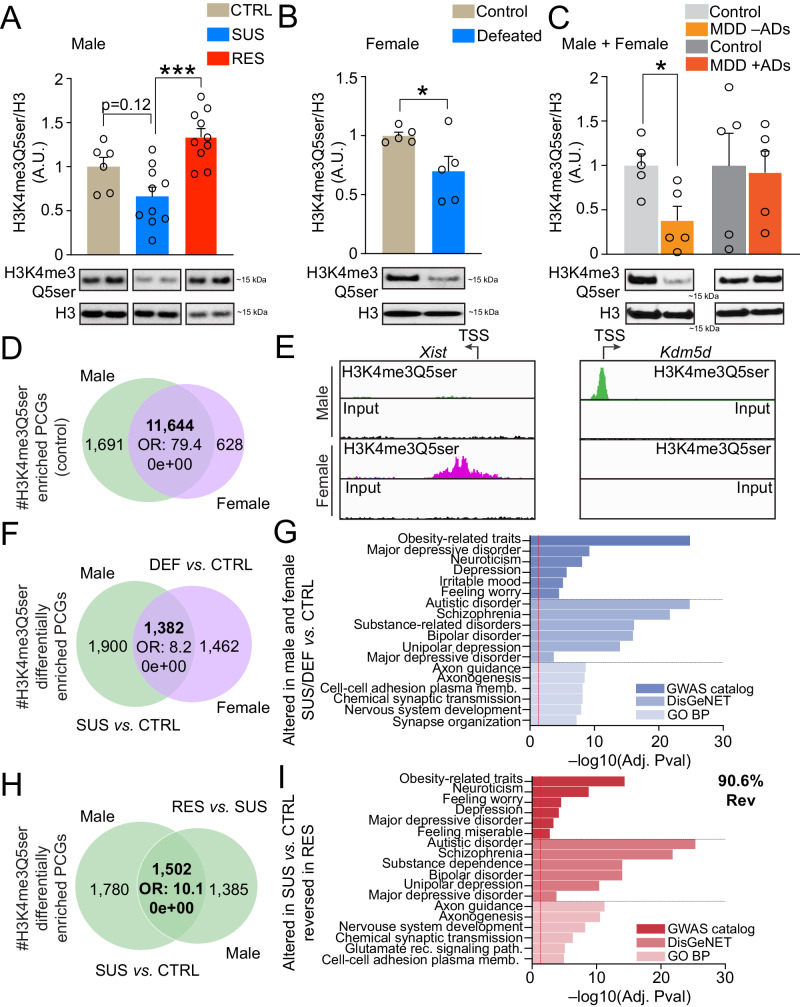
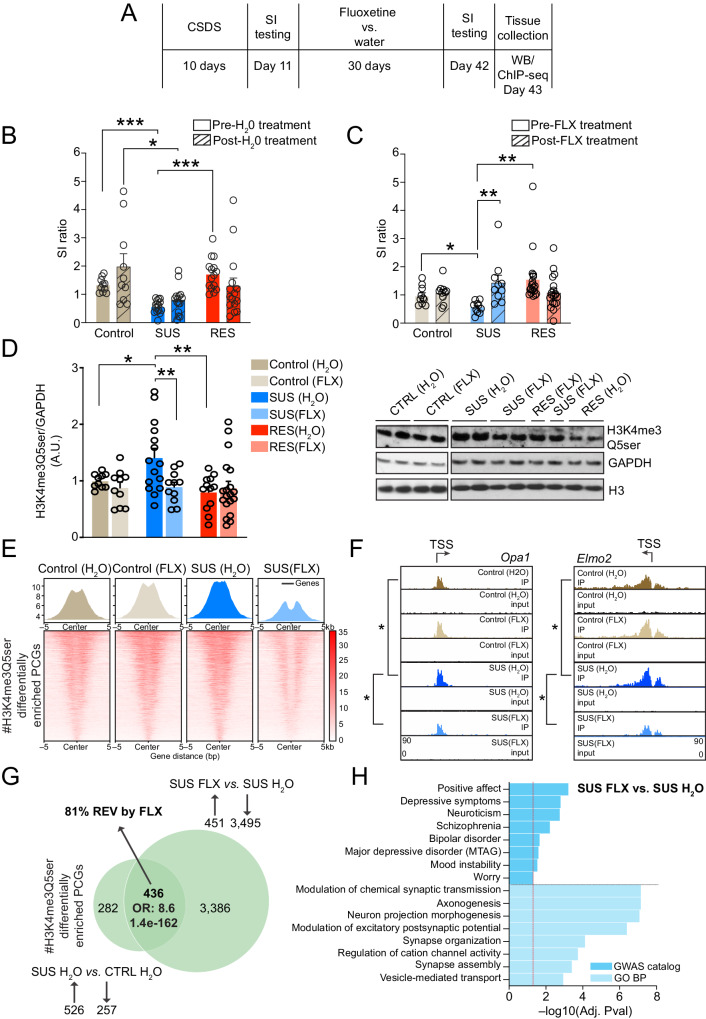
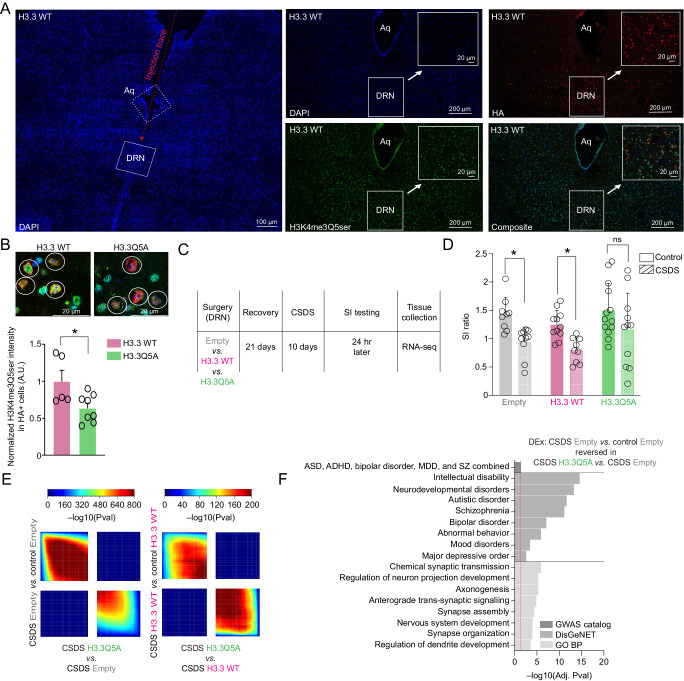
Mood disorders are an enigmatic class of debilitating illnesses that affect millions of individuals worldwide. While chronic stress clearly increases incidence levels of mood disorders, including major depressive disorder (MDD), stress-mediated disruptions in brain function that precipitate these illnesses remain largely elusive. Serotonin-associated antidepressants (ADs) remain the first line of therapy for many with depressive symptoms, yet low remission rates and delays between treatment and symptomatic alleviation have prompted skepticism regarding direct roles for serotonin in the precipitation and treatment of affective disorders. Our group recently demonstrated that serotonin epigenetically modifies histone proteins (H3K4me3Q5ser) to regulate transcriptional permissiveness in brain. However, this non-canonical phenomenon has not yet been explored following stress and/or AD exposures. Here, we employed a combination of genome-wide and biochemical analyses in dorsal raphe nucleus (DRN) of male and female mice exposed to chronic social defeat stress, as well as in DRN of human MDD patients, to examine the impact of stress exposures/MDD diagnosis on H3K4me3Q5ser dynamics, as well as associations between the mark and depression-related gene expression. We additionally assessed stress-induced/MDD-associated regulation of H3K4me3Q5ser following AD exposures, and employed viral-mediated gene therapy in mice to reduce H3K4me3Q5ser levels in DRN and examine its impact on stress-associated gene expression and behavior. We found that H3K4me3Q5ser plays important roles in stress-mediated transcriptional plasticity. Chronically stressed mice displayed dysregulated H3K4me3Q5ser dynamics in DRN, with both AD- and viral-mediated disruption of these dynamics proving sufficient to attenuate stress-mediated gene expression and behavior. Corresponding patterns of H3K4me3Q5ser regulation were observed in MDD subjects on vs. off ADs at their time of death. These findings thus establish a neurotransmission-independent role for serotonin in stress-/AD-associated transcriptional and behavioral plasticity, observations of which may be of clinical relevance to human MDD and its treatment.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Histone H3 serotonylation dynamics in dorsal raphe nucleus contribute to stress- and antidepressant-mediated gene expression and behavior.bioRxiv [Preprint]. 2023 Sep 16:2023.05.04.539464. doi: 10.1101/2023.05.04.539464. bioRxiv. 2023. Update in: Nat Commun. 2024 Jun 13;15(1):5042. doi: 10.1038/s41467-024-49336-4. PMID: 37205414 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
