

Tissue-intrinsic beta-catenin signals antagonize Nodal-driven anterior visceral endoderm differentiation
- PMID: 38871742
- PMCID: PMC11176336
- DOI: 10.1038/s41467-024-49380-0
Tissue-intrinsic beta-catenin signals antagonize Nodal-driven anterior visceral endoderm differentiation
Abstract
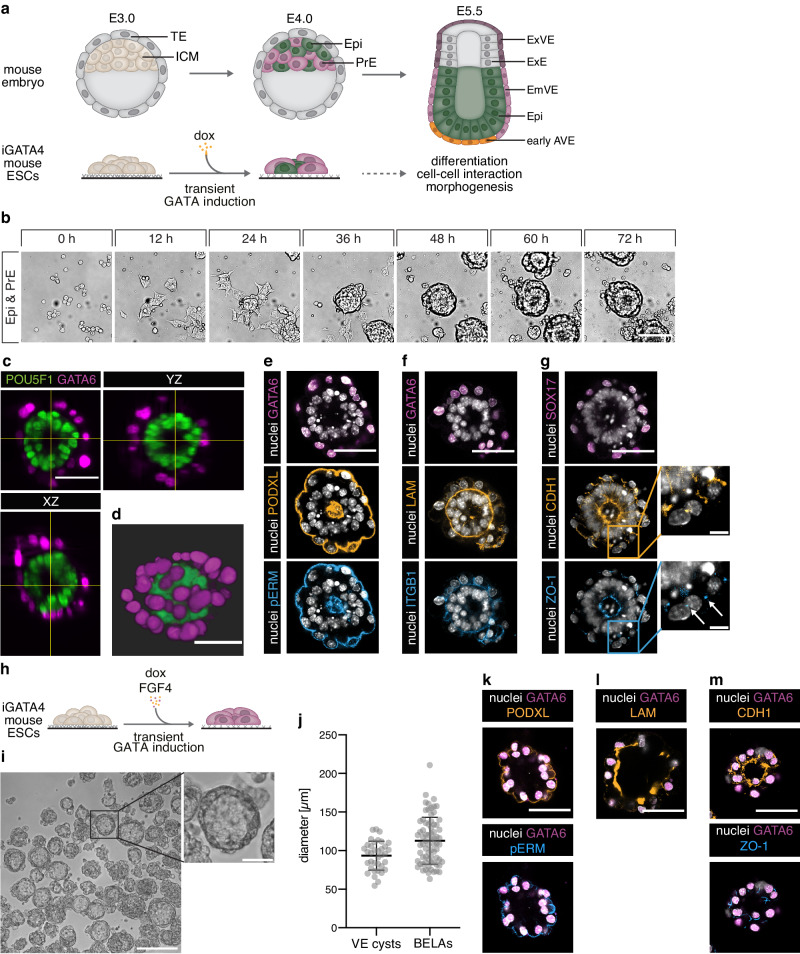
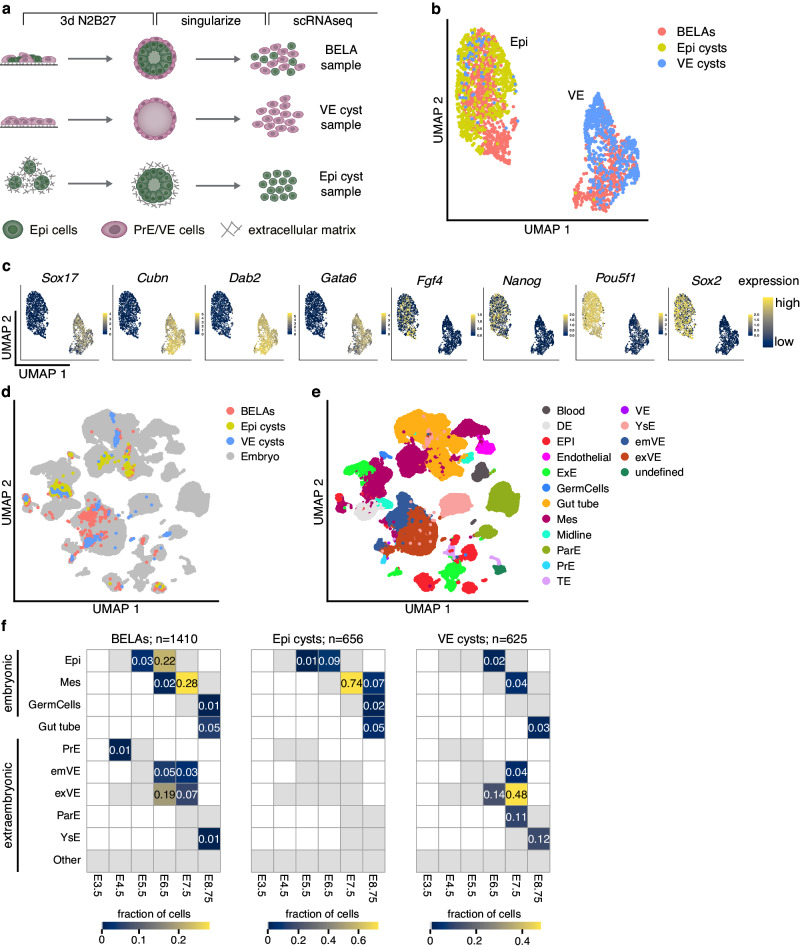
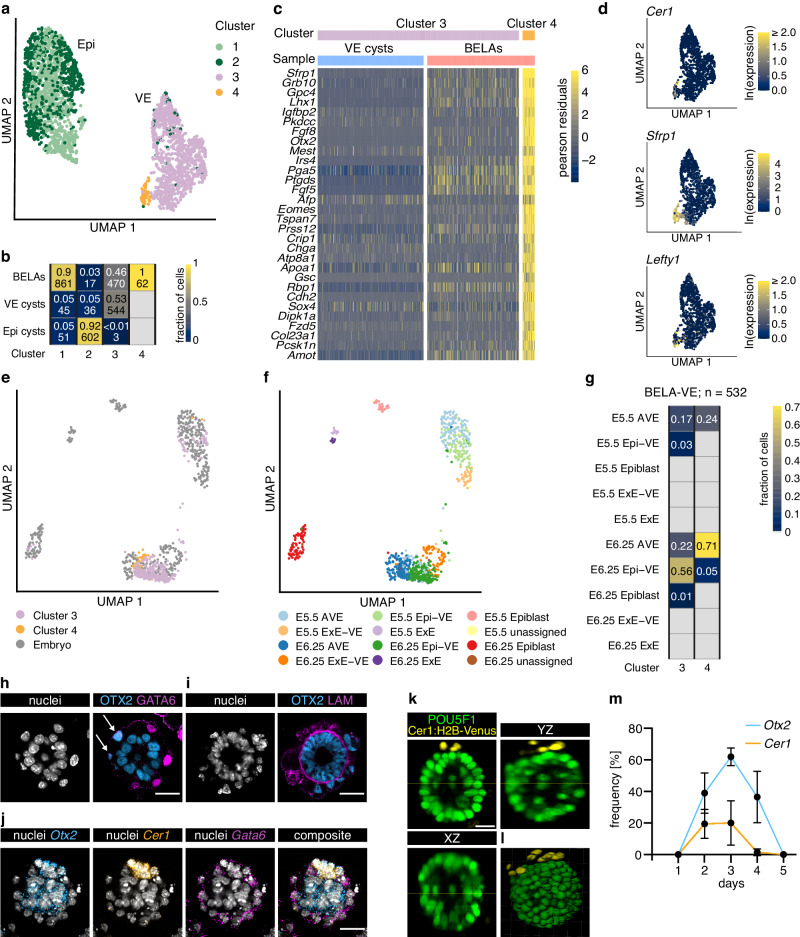
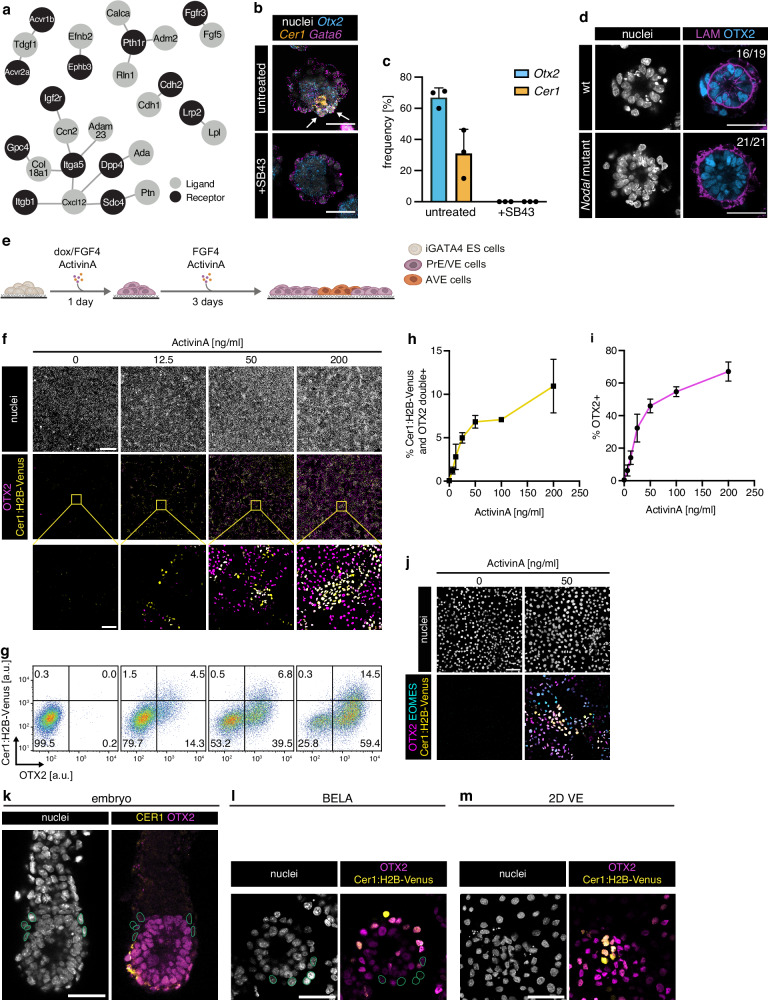
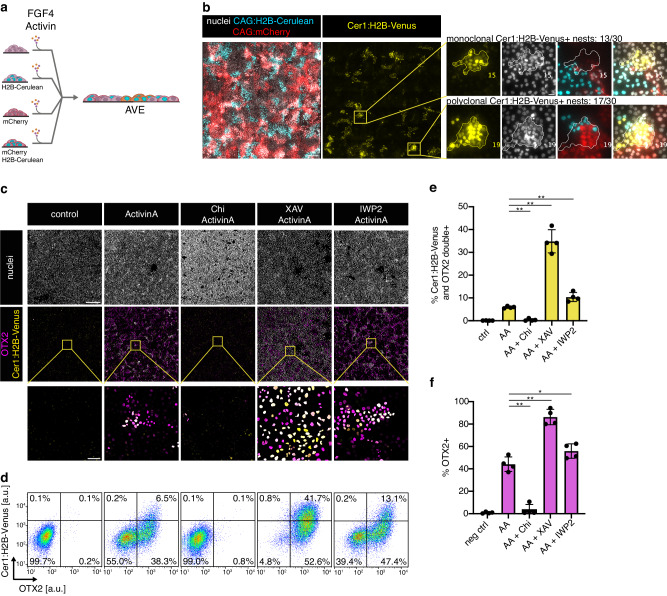
The anterior-posterior axis of the mammalian embryo is laid down by the anterior visceral endoderm (AVE), an extraembryonic signaling center that is specified within the visceral endoderm. Current models posit that AVE differentiation is promoted globally by epiblast-derived Nodal signals, and spatially restricted by a BMP gradient established by the extraembryonic ectoderm. Here, we report spatially restricted AVE differentiation in bilayered embryo-like aggregates made from mouse embryonic stem cells that lack an extraembryonic ectoderm. Notably, clusters of AVE cells also form in pure visceral endoderm cultures upon activation of Nodal signaling, indicating that tissue-intrinsic factors can restrict AVE differentiation. We identify β-catenin activity as a tissue-intrinsic factor that antagonizes AVE-inducing Nodal signals. Together, our results show how an AVE-like population can arise through interactions between epiblast and visceral endoderm alone. This mechanism may be a flexible solution for axis patterning in a wide range of embryo geometries, and provide robustness to axis patterning when coupled with signal gradients.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Nodal signaling from the visceral endoderm is required to maintain Nodal gene expression in the epiblast and drive DVE/AVE migration.Dev Biol. 2015 Apr 1;400(1):1-9. doi: 10.1016/j.ydbio.2014.12.016. Epub 2014 Dec 20. Dev Biol. 2015. PMID: 25536399 Free PMC article.
-
Regionalised signalling within the extraembryonic ectoderm regulates anterior visceral endoderm positioning in the mouse embryo.Mech Dev. 2006 Apr;123(4):288-96. doi: 10.1016/j.mod.2006.01.004. Epub 2006 Mar 3. Mech Dev. 2006. PMID: 16517131
-
Regulation of extra-embryonic endoderm stem cell differentiation by Nodal and Cripto signaling.Development. 2011 Sep;138(18):3885-95. doi: 10.1242/dev.065656. Development. 2011. PMID: 21862554 Free PMC article.
-
Dose-dependent Nodal/Smad signals pattern the early mouse embryo.Semin Cell Dev Biol. 2014 Aug;32:73-9. doi: 10.1016/j.semcdb.2014.03.028. Epub 2014 Apr 1. Semin Cell Dev Biol. 2014. PMID: 24704361 Review.
-
Heading forwards: anterior visceral endoderm migration in patterning the mouse embryo.Philos Trans R Soc Lond B Biol Sci. 2014 Dec 5;369(1657):20130546. doi: 10.1098/rstb.2013.0546. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 25349454 Free PMC article. Review.
Cited by
-
Double assurance in the induction of axial development by egg dorsal determinants in Xenopus embryos.Proc Natl Acad Sci U S A. 2025 Feb 18;122(7):e2421772122. doi: 10.1073/pnas.2421772122. Epub 2025 Feb 10. Proc Natl Acad Sci U S A. 2025. PMID: 39928870 Free PMC article.
-
Med12 cooperates with multiple differentiation signals to facilitate efficient lineage transitions in embryonic stem cells.J Cell Sci. 2025 Apr 15;138(8):jcs263794. doi: 10.1242/jcs.263794. Epub 2025 Apr 30. J Cell Sci. 2025. PMID: 40237177 Free PMC article.
References
MeSH terms
Substances
Grants and funding
- A130140/Volkswagen Foundation (VolkswagenStiftung)
- 441798639/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 1328-567/Federal Ministry of Food and Agriculture | Bundesinstitut für Risikobewertung (Federal Institute for Risk Assessment)
- 101043753/EC | EC Seventh Framework Programm | FP7 Ideas: European Research Council (FP7-IDEAS-ERC - Specific Programme: "Ideas" Implementing the Seventh Framework Programme of the European Community for Research, Technological Development and Demonstration Activities (2007 to 2013))
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
