

Partitioning into ER membrane microdomains impacts autophagic protein turnover during cellular aging
- PMID: 38871812
- PMCID: PMC11176346
- DOI: 10.1038/s41598-024-64493-8
Partitioning into ER membrane microdomains impacts autophagic protein turnover during cellular aging
Abstract
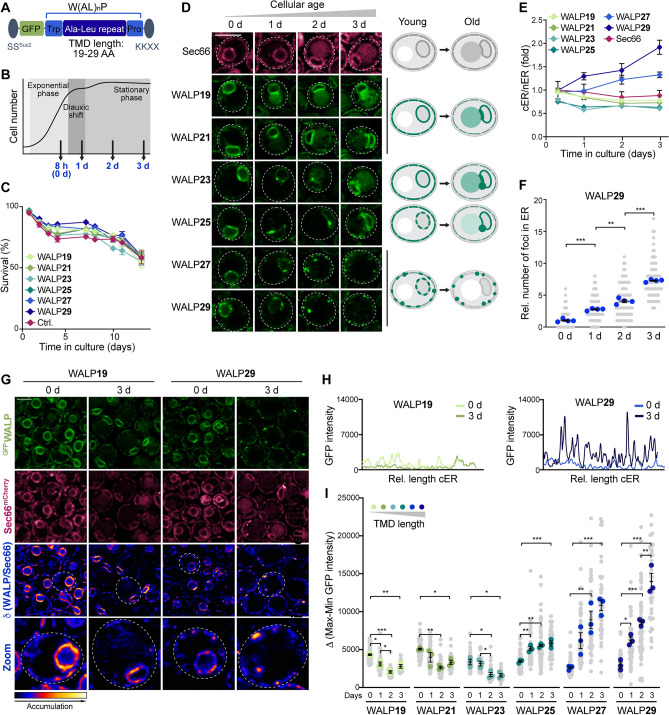
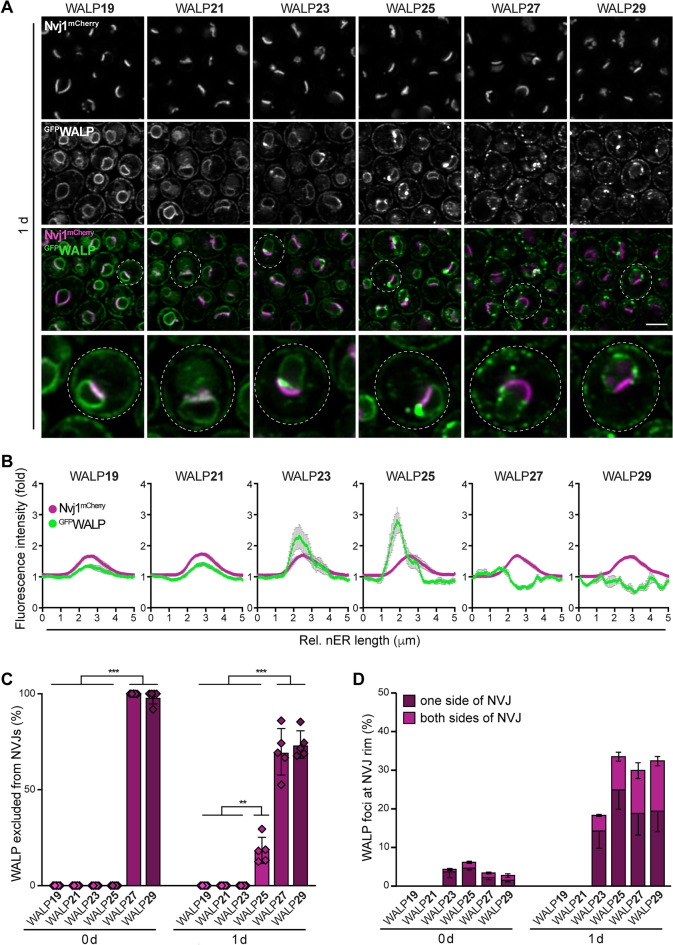
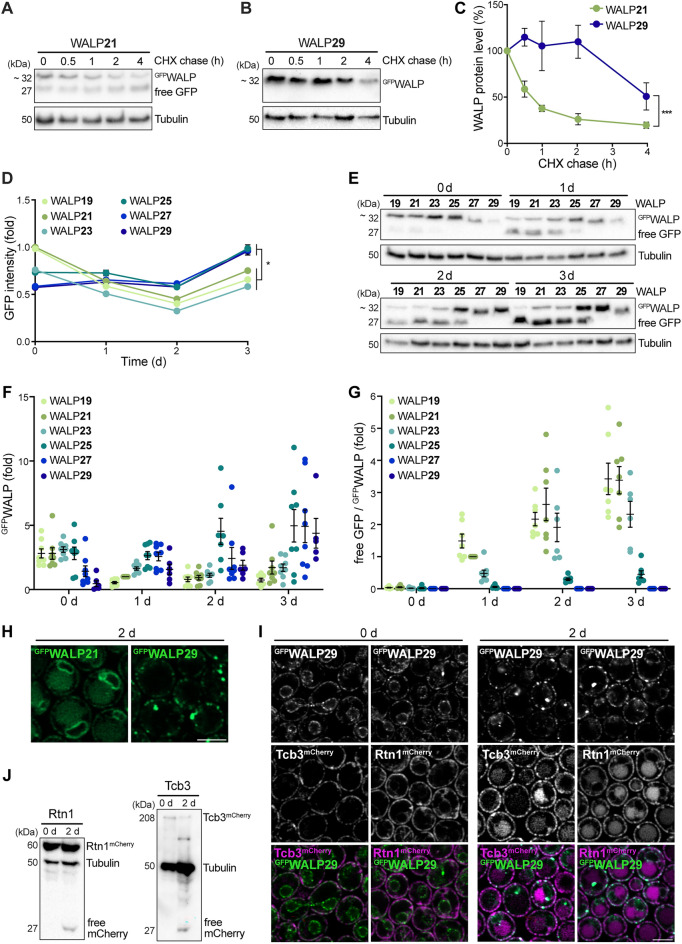
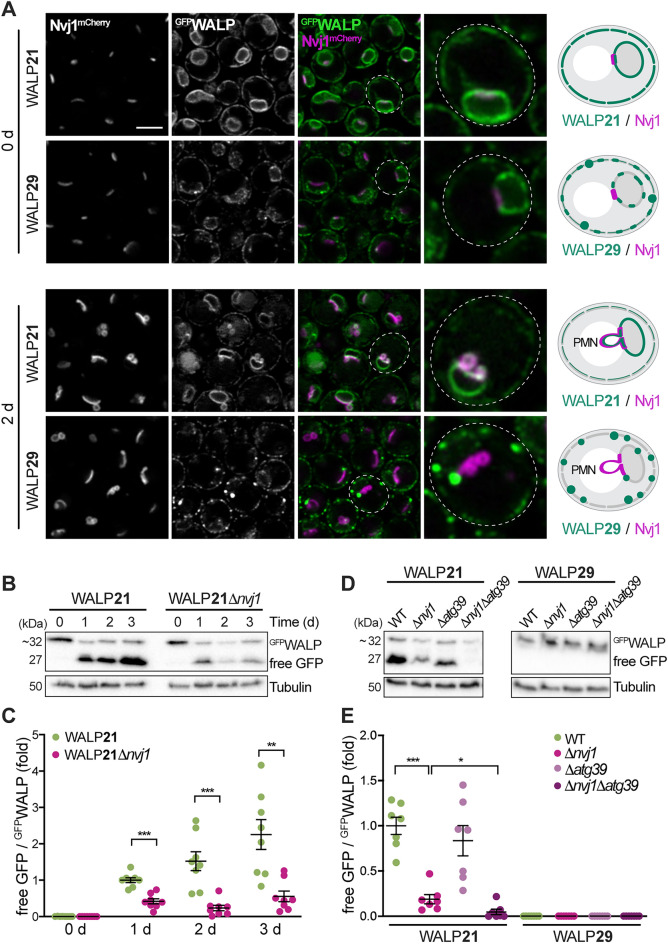
Eukaryotic membranes are compartmentalized into distinct micro- and nanodomains that rearrange dynamically in response to external and internal cues. This lateral heterogeneity of the lipid bilayer and associated clustering of distinct membrane proteins contribute to the spatial organization of numerous cellular processes. Here, we show that membrane microdomains within the endoplasmic reticulum (ER) of yeast cells are reorganized during metabolic reprogramming and aging. Using biosensors with varying transmembrane domain length to map lipid bilayer thickness, we demonstrate that in young cells, microdomains of increased thickness mainly exist within the nuclear ER, while progressing cellular age drives the formation of numerous microdomains specifically in the cortical ER. Partitioning of biosensors with long transmembrane domains into these microdomains increased protein stability and prevented autophagic removal. In contrast, reporters with short transmembrane domains progressively accumulated at the membrane contact site between the nuclear ER and the vacuole, the so-called nucleus-vacuole junction (NVJ), and were subjected to turnover via selective microautophagy occurring specifically at these sites. Reporters with long transmembrane domains were excluded from the NVJ. Our data reveal age-dependent rearrangement of the lateral organization of the ER and establish transmembrane domain length as a determinant of membrane contact site localization and autophagic degradation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Lipid Droplets and Their Autophagic Turnover via the Raft-Like Vacuolar Microdomains.Int J Mol Sci. 2021 Jul 29;22(15):8144. doi: 10.3390/ijms22158144. Int J Mol Sci. 2021. PMID: 34360917 Free PMC article. Review.
-
ER-phagy mediates selective degradation of endoplasmic reticulum independently of the core autophagy machinery.J Cell Sci. 2014 Sep 15;127(Pt 18):4078-88. doi: 10.1242/jcs.154716. Epub 2014 Jul 22. J Cell Sci. 2014. PMID: 25052096 Free PMC article.
-
Lipid droplet biogenesis is spatially coordinated at ER-vacuole contacts under nutritional stress.EMBO Rep. 2018 Jan;19(1):57-72. doi: 10.15252/embr.201744815. Epub 2017 Nov 16. EMBO Rep. 2018. PMID: 29146766 Free PMC article.
-
Roles for L o microdomains and ESCRT in ER stress-induced lipid droplet microautophagy in budding yeast.Mol Biol Cell. 2021 Dec 1;32(22):br12. doi: 10.1091/mbc.E21-04-0179. Epub 2021 Oct 20. Mol Biol Cell. 2021. PMID: 34668753 Free PMC article.
-
Autophagic degradation of the endoplasmic reticulum.Proc Jpn Acad Ser B Phys Biol Sci. 2020;96(1):1-9. doi: 10.2183/pjab.96.001. Proc Jpn Acad Ser B Phys Biol Sci. 2020. PMID: 31932525 Free PMC article. Review.
Cited by
-
Role of membrane microdomains in cardiac protection: strategies for diabetic cardiomyopathy.Am J Physiol Heart Circ Physiol. 2025 Aug 1;329(2):H572-H591. doi: 10.1152/ajpheart.00139.2025. Epub 2025 Jul 10. Am J Physiol Heart Circ Physiol. 2025. PMID: 40637400 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
