

Chilling stress response in tobacco seedlings: insights from transcriptome, proteome, and phosphoproteome analyses
- PMID: 38872895
- PMCID: PMC11170286
- DOI: 10.3389/fpls.2024.1390993
Chilling stress response in tobacco seedlings: insights from transcriptome, proteome, and phosphoproteome analyses
Abstract
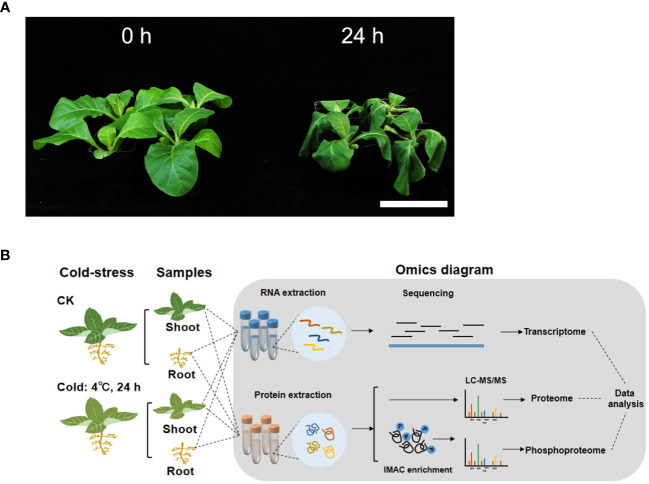
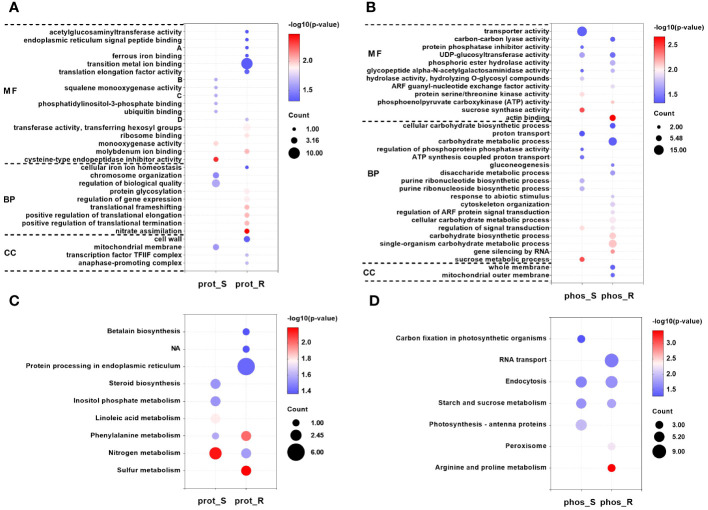
Tobacco (Nicotiana tabacum L.) is an important industrial crop, which is sensitive to chilling stress. Tobacco seedlings that have been subjected to chilling stress readily flower early, which seriously affects the yield and quality of their leaves. Currently, there has been progress in elucidating the molecular mechanisms by which tobacco responds to chilling stress. However, little is known about the phosphorylation that is mediated by chilling. In this study, the transcriptome, proteome and phosphoproteome were analyzed to elucidate the mechanisms of the responses of tobacco shoot and root to chilling stress (4 °C for 24 h). A total of 6,113 differentially expressed genes (DEGs), 153 differentially expressed proteins (DEPs) and 345 differential phosphopeptides were identified in the shoot, and the corresponding numbers in the root were 6,394, 212 and 404, respectively. This study showed that the tobacco seedlings to 24 h of chilling stress primarily responded to this phenomenon by altering their levels of phosphopeptide abundance. Kyoto Encyclopedia of Genes and Genomes analyses revealed that starch and sucrose metabolism and endocytosis were the common pathways in the shoot and root at these levels. In addition, the differential phosphopeptide corresponding proteins were also significantly enriched in the pathways of photosynthesis-antenna proteins and carbon fixation in photosynthetic organisms in the shoot and arginine and proline metabolism, peroxisome and RNA transport in the root. These results suggest that phosphoproteins in these pathways play important roles in the response to chilling stress. Moreover, kinases and transcription factors (TFs) that respond to chilling at the levels of phosphorylation are also crucial for resistance to chilling in tobacco seedlings. The phosphorylation or dephosphorylation of kinases, such as CDPKs and RLKs; and TFs, including VIP1-like, ABI5-like protein 2, TCP7-like, WRKY 6-like, MYC2-like and CAMTA7 among others, may play essential roles in the transduction of tobacco chilling signal and the transcriptional regulation of the genes that respond to chilling stress. Taken together, these findings provide new insights into the molecular mechanisms and regulatory networks of the responses of tobacco to chilling stress.
Keywords: chilling stress; molecular mechanism; phosphoproteome; proteome; tobacco; transcriptome.
Copyright © 2024 Shao, Zhang, Yang, Yu, Guo, Li, Xu and Pan.
Conflict of interest statement
FY, YY and JG were employed by China National Tobacco Corporation, Guangdong Company. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
Physiological and ecological responses of flue-cured tobacco to field chilling stress: insights from metabolomics and proteomics.Front Plant Sci. 2024 Nov 25;15:1490633. doi: 10.3389/fpls.2024.1490633. eCollection 2024. Front Plant Sci. 2024. PMID: 39670264 Free PMC article.
-
Differential transcriptome profiling of chilling stress response between shoots and rhizomes of Oryza longistaminata using RNA sequencing.PLoS One. 2017 Nov 30;12(11):e0188625. doi: 10.1371/journal.pone.0188625. eCollection 2017. PLoS One. 2017. PMID: 29190752 Free PMC article.
-
Integrated transcriptomics and metabolomics provides insights into the Nicotiana tabacum response to heat stress.Front Plant Sci. 2024 Jul 22;15:1425944. doi: 10.3389/fpls.2024.1425944. eCollection 2024. Front Plant Sci. 2024. PMID: 39109058 Free PMC article.
-
Transcriptome Profiling, Biochemical and Physiological Analyses Provide New Insights towards Drought Tolerance in Nicotiana tabacum L.Genes (Basel). 2019 Dec 15;10(12):1041. doi: 10.3390/genes10121041. Genes (Basel). 2019. PMID: 31847498 Free PMC article.
-
Integrative Proteome and Phosphoproteome Profiling of Early Cold Response in Maize Seedlings.Int J Mol Sci. 2022 Jun 10;23(12):6493. doi: 10.3390/ijms23126493. Int J Mol Sci. 2022. PMID: 35742945 Free PMC article.
Cited by
-
Physiological and ecological responses of flue-cured tobacco to field chilling stress: insights from metabolomics and proteomics.Front Plant Sci. 2024 Nov 25;15:1490633. doi: 10.3389/fpls.2024.1490633. eCollection 2024. Front Plant Sci. 2024. PMID: 39670264 Free PMC article.
References
-
- Almadanim M. C., Alexandre B. M., Rosa M. T. G., Sapeta H., Leitão A. E., Ramalho J. C., et al. . (2017). Rice calcium-dependent protein kinase OsCPK17 targets plasma membrane intrinsic protein and sucrose-phosphate synthase and is required for a proper cold stress response. Plant Cell Environ. 40, 1197–1213. doi: 10.1111/pce.12916 - DOI - PubMed
LinkOut - more resources
Full Text Sources